Zur Kenntnis der Spicularzellen und Calciumoxalatidioblasten, sowie der Blattanatomie der Welwitschia.
Inaugural-Dissertation zur Erlangung der Doktorwürde der hohen Philosophischen Fakultät der Universität Rostock
vorgelegt von Curt Bleisch aus Strehlen.
Erler's Buchdruckerei in 1891.
Referent: Prof. Dr. Falkenberg.
Seinen teuren Eltern in kindlicher Liebe gewidmet
vom Verfasser.
Einleitung.
Über das allgemeine Verhalten, sowie die Entstellung und physiologische Bedeutung des Calciumoxalats sind verschiedene Ansichten geäussert worden:
Mulder [1] denkt sich die Oxalsäure durch Desoxydation der Kohlensäure entstanden.
Schleiden [2] hält sie für ein im Stoffwechsel notwendig erzeugtes Produkt, welches auf den Pflanzenorganismus giftig wirkt und durch Bindung an Kalk für die Pflanze unschädlich gemacht wird.
Holzner [3] glaubt, dass die Oxalsäure aus Protëinstoffen gebildet wird, und hauptsächlich dazu bestimmt sei, die aufgelösten Schwefel- und phosphorsauren Kalksalze, welche aus dem Boden aufgenommen werden, an zersetzen und die Säuren frei zu machen, um den für das Fortbestehen der Pflanze notwendigen Phosphor und Schwefel zu liefern.
Pfitzer [4], welcher sich mit den Einschlüssen von Calciumoxalat in die pflanzliche Zellmembran beschäftigt hat, ist der Ansicht, dass dieser Einschluss in zweierlei Art bewirkt werden kann, "entweder durch eine Art Überwallung, indem die Membran um den Krystall sich erhebt und über ihm verschmilzt, oder indem derselbe vom Plasma mit Zellstoff überdeckt wird."
De Vries [5] fasst die Resultate seiner Untersuchungen in folgendem zusammen: "Die Oxalsäure entsteht, wie die Pflanzensäuren wahrscheinlich im allgemeinen, in den turgescirenden Zellen, bei höheren Pflanzen also im Parenchym, und verbindet sich sofort zum Theil oder ganz mit, dem im Zellsaft gelösten Kalk zu schwer oder auch ganz unlöslichem Calciumoxalat, Ist die Lösung dieses letzteren Salzes gesättigt, so krystallisiert es an morphologisch dazu bestimmten Orten aus. Diese letzteren fügen sich der Kegel, dass dieser Auswurfstoff (Calciumoxalat) dem Stoffwechsel möglichst entzogen, und dort abgelagert wird, wo er diesen am wenigsten beeinträchtigt."
Einen erfolgreichen Schritt in der Lösung der Calciumoxalatfrage haben Schimper [6] und Kohl [7] durch ihre bedeutsamen Arbeiten gethan. Ersterer spricht in seiner Einleitung die Ansicht aus, dass der Bildung des oxalsauren Kalks nicht stets dieselbe Bedeutung für den Stoffwechsel zuzuschreiben sei. Dieser Gesichtspunkt war in den vorher genannten Arbeiten vollständig ausser Acht gelassen worden. Schimper unterscheidet nach der Art der Entstehung drei Arten Calciumoxalat:
- primäres, welches sich während des Blattwachsthums unabhängig vom Licht bildet,
- secundäres, welches unter Einfluss von Licht im ausgewachsenen Blatt erzeugt wird,
- tertiäres, welches bei der herbstlichen Entleerung der Blätter gebildet wird.
Ferner fasst er die Aufgabe des Kalks als eine zweifache auf. Erstens führt er der Pflanze Stickstoff, Phosphor und Schwefel in Form von Säuren zu. Zweitens dient er aber auch zum Transport der Kohlenhydrate, da bei Kalkmangel die Stärke in den assimilirenden Zellen angehäuft wird, während die leitenden Zellen stärkearm bleiben. Kohl bestätigt in seiner umfangreichen Arbeit die Resultate Schimpers. Er nimmt aber zum Unterschiede von jenem eine vierte Art Calciumoxalat an, welches er tertiäres nennt, während er das bei der herbstlichen Entleerung entstehende als quartäres bezeichnet. Tertiäres Calciumoxalat ist für ihn dasjenige, welches bei grösseren Cellulosemassen entsteht. Auf Grund der von ihm constatirten Thatsachen spricht er die Ansicht aus, dass die bei der Assimilation gebildeten Kohlenhydrate in Form einer Glycose-Kalk-Verbindung wanderungsfähig seien. Es bildet sich deshalb überall, wo diese Kohlenhydrate in irgend welcher Form, Stärke oder Cellulose, sich ausscheiden, durch die hinzutretende Oxalsäure Calciumoxalat. Zu denselben Untersuchungsergebnissen kommt auch Alberto Alberti [8].
Schimper [9] glaubt in Folge seiner neuesten Arbeiten betonen zu müssen, dass man entgegen diesen jetzt wohl allgemeinen Ansichten über die Bedeutung des Calciumoxalats im Zusammenhang mit den wandernden Kohlenhydraten demselben doch keine solche bedeutsame Rolle im Stoffwechsel einräumen dürfe. Ich selbst hatte meine Arbeiten beim Erscheinen dieser neuesten Schimper'schen Arbeit bereits abgeschlossen, und habe eine Nachuntersuchung seiner Angaben nicht vorgenommen. Deshalb beschränke ich mich auf ein kurzes Referat der einschlägigen Abschnitte der Schimper'schen Abhandlung. Er nimmt erstens an, dass "die Vorgänge, welche der primären, secundären und tertiären Calciumoxalatbildung entsprechen, auch in calciumoxalatfreien Pflanzen vor sich gehen können, mit dem Unterschiede, dass die Oxalsäure durch andere organische Säuren ersetzt wird. Zu diesen letzteren zählt er im besonderen die Apfel- und die Weinsäure. Zweitens widerruft er die in seiner ersten Arbeit ausgesprochene und mit vielen anderen Forschern bisher geteilte, Ansicht, dass der Kalk die in den assimilierenden Zellen gebildeten Kohlenhydrate transportire, weil das Absterben der Pflanze bei Kalkmangel bei höherer Temperatur unterbleibt, wie Dehérain festgestellt hat, und ferner kalkfrei gezogene Tradescautien in eine 2 % Zuckerlösung gelegt, ein reichliches Eindringen in ihre Gewebe, sowie eine Umwandelung in Glycose zeigten. Es hört nach ihm also die Wanderung der Kohlenhydrate nicht vollständig auf bei Kalkmangel, wenn sie auch stark beeinträchtigt wird. Während er dem Kalium und Magnesium eine überaus wichtige Rolle im Stoffwechsel zuschreibt, betrachtet er den Kalk nur als Vehikel für die Mineralsäuren; der oxalsaure Kalk entsteht nach ihm durch Wechselzersetzung der Kalksalze der Mineralsäuren mit den Kalioxalaten, welchen Prozess er für notwendig hält, da durch Anhäufung dieses und spec. des sauren Salzes pathologische Erscheinungen an der Pflanze hervorgerufen werden. Er greift mit dieser Ansicht auf ältere Anschauungen, z. B. von de Vries zurück.
In wie weit diese Ansichten richtig und allgemein gültig sind, lasse ich dahingestellt. Jedenfalls wird durch die vielfach erscheinenden neuen Arbeiten über das Calciumoxalat, welche immer neue Gesichtspunkte zu Tage fördern und die mehr oder weniger wichtige Rolle dieses Salzes für den Stoffwechsel immer mannigfaltiger erscheinen lassen, die Frage ihrer Lösung entgegengeführt werden.
Was nun insbesondere das Vorkommen des Calciumoxalats in den Idioblasten betrifft, so finden wir die erste darauf bezügliche Beobachtung in J. D. Hooker's [10] berühmter Abhandlung über Welwitschia mirabilis. Er fand hier im Blatt sowohl wie im Stamm mehr oder weniger verästete, stark verdickte Zellen, welche mit Krystallen bedeckt sind. Er nannte sie "Spicularzellen". Mit der feineren Structur, ihrem chemischen Charakter, sowie ihrer Bedeutung für die Pflanze hat er sich nicht beschäftigt. Dr. Frankland, welchem J. D. Hooker die Untersuchung der Krystalle der Spicularzellen übergab, hielt dieselben für kieselsäurehaltig.
Colonel Philipp Yorke [11] erkannte zwar, dass die Krystalle nach dem Glühen die Reactionen von kohlensaurem Kalk zeigten, konnte sich jedoch ebenfalls kein bestimmtes Urtheil bilden, weil er sich ihre scheinbare Unlöslichkeit in Salzsäure und Fluorwasserstoff nicht zu erklären wusste. Er nahm an, dass die Krystalle in eine sie vor der Einwirkung der Säuren schützende Substanz eingehüllt, seien, Jedenfalls rührte der Irrtum dieses Forschers, wie schon Graf Solms [12] ausgesprochen hat, davon her, dass er die zurückgebliebenen Löcher nach der Lösung für die Krystalle selbst hielt Hofmeister [13] endlich charakterisierte die Krystalle durch ihr Verhalten gegen Salzsäure und Essigsäure als oxal-sauren Kalk.
Späterhin wurden ähnliche Zellen in Stamm und Blatt einiger Coniferen (Dammara, Araucaria, Sciadopitys) entdeckt und von Mohl [14] zuerst erwähnt, Ebenso erkannte derselbe, dass die früher als "Cuticularknoten" gedeuteten Höcker der verzweigten Sternhaare in den Intercellulargängen von Nymphacaceen Calciumoxalatkrystalle seien. Sie müssen deshalb den Krystallidioblasten nahe stehen.
Da die Arbeiten über die Krystallidioblasten grösstenteils älteren Datums sind, so war wohl anzunehmen, daß eine erneute Untersuchung über diesen Gegenstand manches klarer stellen würde unter Benutzung der neueren mikroskopischen Technik, insbesondere derjenigen Farbstoffe und Reagentien, welche erst in neuerer Zeit von der Industrie dargestellt, wertvolle Hülfsmittel geworden sind, chemisch verschieden zusammengesetzte Elemente des Pflanzenorganismus durch characteristische Farbenreactionen scharf von einander zu trennen.
Ich habe mir deshalb die Aufgabe gestellt, die Calciumoxalatidioblasten mikrochemisch zu untersuchen, ihre Entstellung und Entwicklung zu verfolgen und aus den Resultaten der Untersuchung Schlüsse für ihre physiologische Bedeutung zu ziehen.
Die Arbeit selbst teile ich in drei Abschnitte: Das Calciumoxalat in den Idioblasten
- des Blattes von Welwitschia mirabilis,
- der Coniferen,
- der Nymphaeaceen.
Meine Untersuchungen wurden im pflanzenphysiologischen Institut der Universität Breslau ausgeführt. Anregung hierzu erhielt ich von Herrn Geheimen Regierungsrat Prof. Dr. Cohn. Es möge mir an dieser Stelle vergönnt sein, meinem hochver-ehrten Lehrer für seine liebenswürdige Unterstützung bei der Durchführung der Arbeit, sowie für die gütige Überlassung des Materials meinen wärmsten und aufrichtigsten Dank auszusprechen.
Cap. I.
Das Calciumoxalat in den Idioblasten der Blätter v. Welwitschia Mirabilis Hook.
Die beiden Laubblätter der Welwitschia mirabilis Hook. sind flach, lederartig, parallelnervig und auf der Unterseite von hellerer Färbung wie auf der Oberseite. Sie erreichen eine bedeutende Länge und zerschlitzen im Alter bis zur Basis. Da die bisherigen Untersuchungen über die Anatomie des Blattes entweder sehr kurz gefasst oder die einzelnen Angaben sehr zerstreut sind, so erschien es wünschenswert, eine auf eigene Untersuchungen gegründete Darstellung der gesammten Anatomie vorauszuschicken, in welcher auch die älteren darauf bezüglichen Angaben kritisch zusammengefasst werden sollten. Das Material, welches mir zur Untersuchung zu Gebote stand, verdanke ich der Güte des Herrn Geheimen Regierungsrates Prof. Dr. Cohn, welcher mir die von Herrn Privatdocenten Dr. Gürich auf seiner geologischen Expedition nach Damaraland gesammelten und dem hiesigen Museum überwiesenen Exemplare für diesen Zweck überliess.
Der erste, welcher die Anatomie des Welwitschiablattes beschrieb, ist J. D. Hooker [15]. Seine Darstellung aber ist etwas unvollständig und enthält einige Unrichtigkeiten in Bezug auf die Histologie des Innengewebes. Auch den Bau der Spaltöffnungen hat er nur kurz behandelt. Die Abbildungen zur Anatomie des Blattes befinden sich auf Taf. XIV seiner Arbeit. Er giebt hier auch einen Querschnitt desselben, welcher aber in Betreff der Anordnung der einzelnen Elemente nicht genau der Wirklichkeit entspricht und ausserdem falsch orientirt ist, indem das Phloëm des Gefässbündels der Unterseite des Blattes zugekehrt ist, und nicht wie in der gegebenen Zeichnung nach oben liegt, welchen Fehler De Bary in seiner später zu erwähnenden Abhandlung verbesserte.
Eine zweite noch kürzere Abhandlung über diesen Gegenstand giebt Bertrand [16] im Rahmen einer grösseren Untersuchung. Sie bringt nichts neues; die Zeichnungen auf Taf. I u. II sind sehr klein und mangelhaft und stehen den Hooker'schen bei weitem nach.
Eine weitere zusammenfassende Anatomie des Blattes ist in der Literatur nicht vorhanden, doch finden wir zahlreiche Angaben in derselben zerstreut vor.
Graf Solms-Laubach [17] giebt eine kurze Beschreibung der Epidermis des Blattes, in welcher er zuerst eine Körnerzone, deren krystallinischen Charakter er mit Hülfe des Polarisationsapparates nachwies, beobachtete. Er erkannte sie als aus oxalsaurem Kalk bestehend. Eine Zeichnung hierzu findet sich bei dieser Abhandlung auf Taf. VI, Fig. 20. Einige Einzelheiten habe ich jedoch anders beobachtet, besonders auch die Verdickungen der Epidermiszellen und der Schliesszellen der Spaltöffnung. Auch erwähnt er in derselben Abhandlung das Vorkommen des oxalsauren Kalks im Innenparenchym des Blattes.
Tschirch [18] giebt eine kurze Notiz über die Spaltöffnungen des Welwitschiablattes, De Bary [19] berührt in seinem Lehrbuch der vergleichenden Anatomie die anatomischen Verhältnisse des Welwitschiablattes an verschiedenen Stellen seines Buches und giebt im besonderen eine genaue Beschreibung des Gefässbündels, die Zeichnung des Querschnitts (pag. 425) ist zu sehr schematisiert und kann deshalb eine klare Vorstellung von dem natürlichen Bilde nicht geben. Instructiver dagegen ist der Querschnitt eines Hauptgefässbündels (pag. 348), Im allgemeinen haben meine Untersuchungen die Ansichten De Bary's über den Bau des Gefässbündels bestätigt. Weitere kurze Angaben De Bary's über die Anatomie derselben werden am passenden Orte angeführt werden,
Was die Krystallidioblasten oder "Spicularzellen" des Welwitschiablattes betrifft, so hat sich Graf Solms-Laubach [20] eingehend mit diesem Gegenstand beschäftigt. Er findet, dass die Krystalle der äussersten Zone der stark verdickten Zellen eingelagert seien. Die Wandung derselben hält er für verholzte Membransubstanz. Dieser sind die Krystalle, wie er sich ausdrückt, "eingebacken", so dass sie in Form "wellenartig hervortretender meist dunkelgelber Leisten die Krystalle verkittet". Die verholzte Membransubstanz hält er in keiner Weise für quellungsfähig. Ferner nimmt er an, dass jede Spicularzelle von einer aus Cellulose bestehenden und ihr mit den Nachbarzellen gemeinsamen Scheide, einem sogenannten "Saftrohr" umgeben sei. Er kommt zu dem Schluss, "dass die Verdickung und Verholzung, sowie das Vorkommen der Krystalle an die Innenlage der Membran gebunden sei."
Kohl [21] scheint in seiner schon erwähnten grösseren Arbeit über das Calciumoxalat die Verhältnisse desselben bei Welwitschia mirabilis, deren er allerdings Erwähnung thut, nicht untersucht zu haben, da er auf die Graf Solms-Laubach'sche Abhandlung hinweist und nichts wesentliches neues bringt.
Ein Querschnitt durch das Blatt der Welwitschia mirabilis zeigt folgendes Bild: Unter der Epidermis zieht sich das Pallisadengewebe hin auf der Ober- und Unterseite des Blattes, während wir dasselbe bei den Blättern im allgemeinen nur auf ersterer finden. In diesen liegen zerstreut direkt unter der Epidermis starke Bündel von Sclerenchymfasern. Das Pallisadengewebe umschliesst ein aus rundlichen Zellen bestehendes Parenchym, in welchem auch die Gefässbündel verteilt sind. Ferner finden sich in beiden Geweben eigentümlich gestaltete mit Krystallen bedeckte Zellen, die "Hooker'schen Spicularzellen", die cuticula (Fig. I. a) der Epidermis ist stark gewellt, Die darunter liegenden Zellen (Fig. I. c.) sind von länglicher Gestalt und schließen fest und lückenlos aneinander. Sie sind auf der der cuticula zugewendeten Seite bedeutend stärker verdickt als auf den dem Blattinneren zugewandten Wänden, Ihr Lumen zieht sich nach der cuticula in einen engen Hals aus, welcher sich an seinem Ende zu einem kleinen kugelförmigen Hohlraum erweitert. "Die Höhe der Epidermis selbst betragt im Mittel 75 µ.; diejenigen Stellen derselben aber, welche die Spaltöffnungen umschliessen, sind eingesenkt, nur 50 µ. hoch, und bilden in Folge dessen einen die Spaltöffnung umgrenzenden trichterförmigen nach den Schliesszellen zuführenden Vorraum. Die inneren Verdickungen der Epidermiszellen bestehen ans reiner Cellulose; sie färben sich mit Chlorzinkjod blau, mit Congoroth rot, mit Saffranin wenig oder garnicht, während durch letzteres eine tiefrote Färbung des Primordialschlauches hervorgerufen wird. Unter der cuticula bemerken wir in der äusseren Schicht der Epidermiszellen eine körnige Zone (Fig. I b.), welche durch ihr optisches Verhalten als eine krystallinische charakterisirt ist. Die microchemische Untersuchung lässt sie als aus Calciumoxalat bestehend erkennen. Nach der Lösung dieses letzteren in Salz- oder Salpetersäure bleiben an der Stelle der Krystalle schwarze Punkte zurück; das sind die Holzräume der aufgelösten Krystalle. Sie grenzt scharf gegen die eigentliche cuticula, sowie die unter ihr liegenden aus Cellulose bestehenden inneren Verdickungen der Epidermiszellen, welche beide krystallfrei sind, ab, Nach Graf Solms-Laubach [22] und Kohl [23] gehört diese Körnerzone zu den Cuticularschichten, deren unterste sie vorstellt. Sie verhält sich aber nach meinen Beobachtungen wesentlich anders gegen Farbstoffe wie die körnerfreie cuticula, weshalb man sie wohl nicht aufzufassen hat als eine mit der übrigen cuticula chemisch übereinstimmende nur durch den Gehalt an Calciumoxalat verschiedene, sondern als eine auch in ihren chemischen Eigenschaften veränderte Schicht, welche wir wegen ihres Gesammtverhaltens wohl am besten mit dem Namen "Krystallogenschicht" bezeichnen. Als solcher steht sie möglicher Weise der später zu erwähnenden Krystallschicht der Spicularzellen nahe. Das Verhalten der einzelnen Teile der cuticula gegen Farbstoffe war folgendes: Die körnerfreie Cuticula (Fig. I a) wird mit Congorot blassgelb, mit Fuchsin (in gleichen Teilen Glycerin und Wasser gelöst) tiefrot, mit Saffranin ebenfalls rot gefärbt. Die "Krystallogenschicht" giebt mit Congorot eine bräunlich gelbe, mit Fuchsin eine schwachrötliche, mit Saffranin eine braunrote Färbung. Die inneren Schichten der Epidermiszellen werden durch Congorot rot, durch Fuchsin und Saffranin gar nicht gefärbt. In der Epidermis sind die Spaltöffnungen eingesenkt, so zwar, dass von der Oberfläche des Blattes eine becherförmige Grube nach innen führt, welche sich nach dem Grunde zu in einen schmalen Kanal verengert, welcher unmittelbar den Schliesszellen anliegt. Es entsteht auf diese Weise ein weiter Vorhof mit Oberwall, und ein enger kanalartiger Hinterhof mit Unterwall ähnlich wie bei dem Spaltöffnungsapparat von Agave americana. Zuweilen wurde eine Modification dieses Verhältnisses in der Weise beobachtet, daß zwei Spaltöffnungen dicht bei einander liegend bei einem gemeinsamen Oberwall nur durch einen Unterwall von einander getrennt sind. Die cuticula, welche nach den Stellen hin, an denen sie über die Schliesszellen hinweggeht, allmählich einen geringeren Durchmesser annimmt, ist nur durch einen äusserst schwachen Spalt an der Berührung der beiden Schliesszellen unterbrochen. Die abgezogene Epidermis zeigt die Spaltöffnungen zu sechs bis acht in längeren oder kürzeren Parallelreihen, Entsprechend der schon erwähnten doppelten Einsenkung der Epidermis vor den Schliesszellen (Vor- und Hinterhof) werden die Bilder bei höherer oder tieferer Einstellung verschiedene. Bei der ersten Einstellung nämlich sieht man an Stelle der Spaltöffnungen nur hohle fast rechteckige Gruben, die Umrisse des Oberwalls. Bei einer tieferen Einstellung wird die umgebende Oberhaut undeutlicher und es erscheinen in den Gruben zwei in der Mitte auf einander zugebogene Linien, welche als die Ränder des Hinterwalles aufzufassen sind. Bei einer noch tieferen Einstellung endlich wird der Grund des Hinterhofes mit dem Spalt der Schliesszellen sichtbar.
Zum Vergleich wurde nun ein Längsschnitt untersucht. Hierbei erschienen je nach der Lage des ausgeführten Schnittes drei verschiedene Bilder. Erstens lange ununterbrochene Strecken der Epidermis, zweitens solche mit länglichen Einsenkungen, und drittens ebensolche mit Schliesszellen versehene. Diese Verschiedenheiten lassen sich in der Weise erklären, dass das erste Bild einem Schnitt durch die Oberfläche der Epidermis, das zweite einem solchem durch den tiefer gelegenen Vorhof, das dritte einem solchen durch die Schliesszellen selbst entspricht. An Querschnitten konnte auch deutlich in dem unter der Epidermis liegenden aus langgestreckten Zellen bestehenden Pallisadengewebe ein grosser Raum beobachtet werden, welcher durch Auseinanderweichen der Zellen entstanden ist und demnach die Athemhöhle der Spaltöffnung darstellt.
Die Schliesszellen selbst sind sehr eigentümlich gebaut. Hooker [24] spricht sich in seiner Abhandlung nicht bestimmt darüber aus, da er den anatomischen Bau derselben nicht eingehend untersucht hat. Als besonders günstig zum Studium der Schliesszellen erwies sich eine Färbung mit Saffranin, durch welches die cuticula tiefrot, die Epidermiszellen wenig oder garnicht, die verdickten Stellen der Schliesszellen im Gegensatz zu den unverdickten schön rot gefärbt wurden.
Zum leichteren Verständnis soll die an die cuticula stossende Seite der Schliesszellen "Vorderseite" (Fig. II α), die ihr entgegengesetzte dem Innenblatt angewandte "Hinterseite" (Fig. II γ), die an die Epidermiszellen stossende "Rückenseite" (Fig. II β) und endlich diejenige, mit welchen sich die Schliesszellen berühren, "Bauchseite" (Fig. II δ) genannt werden.
Führt man nun auf einem Querschnitt obige Färbung mit Saffranin aus, so entstehen je nach der Höhe des ausgeführten Schnittes sehr verschiedene Bilder, Teils sind die Schliesszellen von ovaler Gestalt und zeigen auf der Vorder- und Hinterseite starke Verdickungen, welche nur wenig Raum zwischen sich lassen und an der Bauchseite zusammenstossend eine schwächere Verdickung bilden, während die Rückenseite davon frei ist. (Fig. II 1.) Bei tieferer Einstellung geht die ovale Form der Schliesszellen in eine mehr längliche über, bei welcher die Verdickungen nicht mehr so stark durch eine schmälere Verdickungsleiste mit einander verbunden sind. (Fig. II 2.) Bei noch tieferer Einstellung endlich verschwinden diese Verdickungsleisten immer mehr, die Verdickungen werden immer schwächer, die Schliesszellen au der Vorder- und Hinterseite immer spitzer, Schliesslich sind die Verdickungen an diesen beiden Seiten der Zellen nur noch in Form eines Doppelhackens sichtbar, welcher von den Enden der Verdickungen beider Schliesszellen gebildet wird. (Fig. II 3).
Auch konnten bei anderen Schnitten die Bilder in der Reihenfolge beobachtet werden, dass die Schliesszellen zuerst länglich mit schmalen Verdickungen erschienen, um erst dann bei tieferer Einstellung in die oben beschriebene Form mit starken Verdickungen überzugehen, Aus diesen Beobachtungen geht hervor, dass die Schliesszellen nicht an allen Stellen eine gleiche Dicke haben, sondern dass vielmehr an der einen Stelle der Durchmesser von der Vorder- nach der Hinterseite dem auf diesem senkrecht in einer Ebene liegenden von der Bauch- nach der Rückenseite ziemlich gleich ist, während an anderen der erstere bedeutend länger als der letztere wird; die Schliesszellen besitzen demnach auf der Vorder- und Hinterseite eine tiefe Einschnürung. Auch die Verdickungen sind an den einzelnen Teilen der Zelle verschieden stark, Auf einem Längsschnitt (Fig. II. 4) finden wir, dass die Form der Schliesszellen ungefähr semmelförmig ist und auf der Vorder- und Hinterseite in der Mitte eine Einschnürung zeigt, an welcher die Verdickungen ausserordentlich stark sind, während sie nach beiden Enden der Zelle zu schwächer werden. Von der Innenseite der Epidermis aus sieht man die beiden Schliesszellen in länglich ovaler Form, oben und unten etwas eingekerbt. Eine Grenze zwischen beiden ist nur undeutlich sichtbar; an den eingekerbten Stellen beobachtet man nach der Mitte zulaufende schwache Verdickungen, Die zwei hellen Flecke an den beiden Seiten der schwachen Grenze der Schliesszellen sind in der Weise zu erklären, dass das durch den Spalt fallende Licht die beiden Hinterseiten der Zellen beleuchtet, Ein solches Bild ist wohl auch die Zeichnung von Hooker [25] (Taf. XIV.) Die beiden hornartigen Fortsätze stellen hierbei die Enden der Verdickungen dar, Bei einer tieferen Einstellung ändert sich das Bild in der Weise, dass der Querdurchmesser der Zellen grösser wird und eine deutliche Grenze zwischen beiden Zellen erscheint, an welcher oben und unten starke Verdickungen sichtbar sind
Schliesslich wird bei noch tieferer Einstellung der Querdurchmesser der Kellen wieder kleiner, die Verdickungen werden undeutlicher, und es erscheint ein deutlicher Spalt,
Aus allen diesen Beobachtungen ergiebt sich nun das Resultat, dass die Gestalt der Schliesszellen in der Weise aufzufassen ist, dass der Querdurchmesser von der Vorder- nach der Hinterseite sich verändert, so zwar, dass er in der Mitte am kürzesten ist und nach beiden Enden der Zelle zu länger wird; dagegen verhält sich der Durchmesser, welchen man sich senkrecht auf dem genannten Durchmesser in einer Ebene von der Bauch- nach der Rückenseite gelegt denkt, umgekehrt. Er ist in der Mitte der Zelle am grössten, und nimmt an beiden Enden derselben an Grosse ab. Die Verdickungen sind in der Weise in den Schliesszellen verteilt, dass die eine auf der Vorderseite, die andere auf der Hinterseite liegt. Beide sind in der Mitte der Zelle an der eingeschnürten Stelle am stärksten und werden nach den Enden verlaufend allmählich schwächer. Die Bauchseite ist wenig, die Rückenseite garnicht verdickt. Fig. III zeigt eine schematische Darstellung dieser Verhältnisse.
Es entsteht nun die Frage, wie diese so eigentümlich gestalteten und verdickten Zellen in ihrer Eigenschaft als Schliesszellen einer Spaltöffnung functioniren. Im folgendem soll versucht werden, den Mechanismus derselben zu erklären. Der Umstand, dass auf der Rückenseite keine Verdickungen sich befinden, ermöglicht eine leichte Communication mit den Nachbarzellen. Befinden sich nun die Schliesszellen im Turgor, so macht sich diese Turgeszenz in der Weise, geltend, dass die an der Vorder- und Hinterseite liegenden nicht dehnbaren Verdickungen passiv parallel aus einander geführt werden. Diese Bewegung hat notwendiger Weise auf die schwach oder garnicht verdickten, mithin dehnbaren Bauch- und Rückenseite den Erfolg, dass sie sich einander nähern, wodurch andererseits ein Auseinanderweichen der Bauchseiten beider Schliesszellen bewirkt wird und ein Spalt entsteht.
Hört der Turgor auf, so nähern sich die Bauchseiten, und der Spalt schliesst sich.
Das unter der Epidermis liegende Pallisadengewebe beisteht aus langgestreckten Zellen, zwischen welchen Intercellulargänge bemerkbar sind. Ihre Membran ist reine Cellulose, sie wird dementsprechend mit Chlorzinkjod blau, mit Congorot rot gefärbt. Mit Saffranin bleibt die Wandung der Zellen ungefärbt, während das contrahirte Plasma eine schön rote Farbe annimmt. Zuweilen wurden in Haufen von unbestimmter Form in den Intercellulargängen dieses Gewebes grosse Mengen von kleinen Krystallen beobachtet, welche durch ihr Verhalten gegen Essig- und Salzsäure als Calciumoxalat bestimmt wurden. Besonders bei jungen Blättern wurde diese Erscheinung beobachtet, während eine Anhäufung in älteren Blättern immer fehlte. Solches Auftreten und Verschwinden von oxalsaurem Kalk wahrend der Entwickelung der Organe wurde auch anderweitig beobachtet. Schimper [26] behauptet, dass derselbe ebenso beweglich sei wie die. Produkte der Assimilation. Die Umsetzung im Stoffwechsel und die Wanderung des Calciumoxalats im Blatt der Welwitschia habe ich nicht weiter untersucht.
Das zwischen dem oberen und unteren Pallisadengewebe liegende Blattparenchym (Fig. V p.) besteht aus rundlichen Zellen, welche einem Collenchym sehr ähnlich sehen, für welches es Hooker [27] (vergl. pag. 11) in seiner Abhandlung irrtümlicher Weise erklärt. Bei genauer Betrachtung und besonders mit Hülfe von polarisirtem Licht erkennt man aber sofort, dass die collenchymatische Natur desselben nur eine scheinbare ist. Denn behandelt man einen Querschnitt mit Salz- oder Salpetersäure, so bleibt ein aus rundlichen Zellen bestehendes Gewebe zurück, dessen Membran sehr wenig gequollen ist. Letztere besteht aus reiner Cellulose. Das collenchymatische Aussehen des Gewebes wird hervorgerufen durch die Einlagerung kleiner Krystalle, welche sich durch ihr optisches Verhalten als oxalsauren Kalk charakterisiren. Graf Solms-Laubach [28] ist der Ansicht, dass diese Krystalle der Mittellamelle der Zellmembran eingelagert seien. Ich konnte weder eine Mittellamelle noch irgend welche Gruben nach dem Auflösen der Krystalle auf den Zellmembranen finden. Für die Annahme, dass die Krystalle nicht einer Schicht der Zellwandung, sondern den Intercellulargängen eingelagert seien, spricht auch der Umstand, dass bei besonders dünnen Schnitten die Krystalle oft von selbst herausfielen.
Im Pallisadengewebe befinden sich unter der Epidermis in kürzeren oder weiteren Zwischenräumen von einander starke Bündel von Sclerenchymfasern. Auch auf der Grenze zwischen Pallisadengewebe und Blattparenchym liegen allerdings kleinere Bündel solcher Fasern,
Eigentümlich ist auch das Vorkommen einzelner Fasern im letzteren Gewebe, in unmittelbarer Nachbarschaft der später zu behandelnden Spicularzellen. In diesem Falle wachsen sie, den ihnen zur Entwickelung gebotenen Baum ausnutzend, in die Intercellulargänge hinein. Sie enthalten kein Calciumoxalat und unterscheiden sich dadurch wesentlich von den Spicularzellen. Ferner sind sie im Gegensatz zu jenen nicht verholzt, sondern bestehen aus reiner Cellulose, Auch sind sie unverzweigt und von langer spindelförmiger Gestalt, Eine Schichtung ist nur undeutlich sichtbar, dagegen wurden sehr deutliche Tüpfelkanäle wahrgenommen. Das Lumen der Zelle ist fast vollständig von den Verdickungen ausgefüllt.
Das Blattparenchym wird durchzogen von einander parallelen Gefässbündeln, welche durch Nebenäste mit einander anastomasiren. Diese Anastomosen endigen zuweilen blind im Parenchym oder treten mit einander in Verbindung. Bei De Bary [29] finden wir eine Abbildung dieser Verhältnisse.
(Pag. 313.) Die Hauptgefässbündel sind von einer Reihe netzfasriger und getüpfelter Zellen umgeben, welche an einzelnen Stellen von Parenchymzellen unterbrochen ist, Diese Zellen sind innhaltsleer und verholzt. Ein gleicher eigentümlicher Zellensaum wurde von Frank [30] im Blatt von Taxus baccata beobachtet, welchen er jedoch zum Gefässbündel rechnet,. Hugo v. Mohl [31] erkannte das allgemeine Vorkommen dieses Saums in der Familie der Coniferen, Er fasst im Gegensatz zu Frank denselben nicht als den Elementen des Gefässbündels ungehörig auf, sondern sucht die Funktion desselben hauptsächlich darin, dass er die Leitungsbahn für den Übertritt des Saftes aus dem Gefässbündel zum Parenchym und von diesem zurück ins Gefässbündel bildet. Dieser Auffassung gemäss nennt er ihn "Transfusionsgewebe". De Bary [32] bezeichnet ihn hingegen als "Tracheidensaum", Er ordnet damit die dieses Gewebe bildenden Zellen unter den anatomischen Begriff von Tracheiden. Zimmermann [33] hält diese Auffassung für unzutreffend und schliesst sich der Ansicht Mohls an, weil er es für zweckmässig halt, nur "die unzweifelhaft für die Durchlüftung sorgenden Organe als Tracheiden zu bezeichnen", So lange die physiologische Aufgabe dieses Gewebes noch nicht genügend aufgeklärt ist, kann man mit Recht den einen oder anderen Namen für dasselbe Gewebe anwenden. An dem eigentlichen Gefässbündel können wir nun wieder einen inneren Teil und zwei denselben halbmondförmig umgebende Sclerenchymbündel unterscheiden. Die stark verdickten Zellen derselben bestehen aus reiner Cellulose. Der innere Teil des Gefässbündels ist eigentümlich gebaut. Der Unterseite des Blattes zugekehrt liegt ein aus engen langgestreckten Zellen bestehendes Gewebe, dessen Membran gequollen ist. Gewöhnlich ist dasselbe vollständig zusammengedrückt und man kann deshalb auf seinem Längsschnitt seine einzelnen Elemente nicht unterscheiden. Es ist auch deshalb nicht möglich, ein sicheres Urteil abzugeben, ob dieses Gewebe Cambiform oder Siebteil des Gefässbündels vorstellt. Am besten können wir es mit dem allgemeinen Namen Phloëm belegen. Der der Oberseite des Blattes zugewendete Teil des Gefässbündels wird gebildet aus weiten lückenlos verbundenen Zellen, deren Membran ausserordentlich stark gequollen ist, besonders in der Nähe der Sclerenchymscheide. Ihre Wandung ist Cellulose. Zwischen diesen Geweben befinden sich fast in der Mitte des ganzen Gefässbündels Tracheen. Doch kommen auch einige versprengt zwischen dem stark gequollenen Gewebe in der Nähe der Sclerenchymscheide vor. Der Umstand, dass die Tracheen fast in der Mitte des ganzen Gefässbündels sich befinden, hat die Veranlassung gegeben, dasselbe als ein bicollaterales aufzufassen. Dagegen aber spricht das Vorkommen einzelner Tracheen zwischen den stark verdickten Zellen, weshalb dieses letztere Gewebe mit der Gesamtheit des centralen Complexes von Tracheen als "Xylem" zu bezeichnen ist. Auf dem Längsschnitt sieht man auf der Seite des Phloëms Netz- und Hoftüpfeltracheen, solche mit Ring- und Spiralverdickungen folgen. Die wenigen Tracheenelemente, welche sich im dickwandigen Gewebe des Xylems befinden, besitzen die letzteren Verdickungen, welche aber sehr verzerrt sind, Bei einer Beobachtung eines Blattstückes, welches mit Schulze'scher Macerationsflüssigkeit behandelt worden war, erwiesen sich die Netz- und Tüpfeltracheen als ächte Gefässe, während die Ring- und Spiraltracheen als Tracheiden bestimmt wurden. De Bary [34] lässt es noch unentschieden, ob diese letzteren den Gefässen oder Tracheiden zuzurechnen seien, und belegt sie mit dem Colectivnamen für beide Elemente "Tracheen".
Was die Anatomie der Nebengefässbündel betrifft, so ist der allgemeine Bau derselben der gleiche wie der der Hauptgefässbündel. Die Tracheen nehmen die Mitte des Bündels ein und man findet hier ebenfalls Netz-, Tüpfel-, Ring- und Spiraltracheen. Das dickwandige Gewebe des Xylems ist jedoch nicht so reichlich ausgebildet und die Wandung desselben nicht so stark gequollen wie bei den Hauptgefässbündeln, Das Transfusionsgewebe ist bis auf einige wenige Netz- und Tüpfeltracheen reduciert. Die Sclerenchymscheiden fehlen vollständig.
Die Spicular- oder Krystallfaserzellen kommen sehr zahlreich, sowohl im Blattparenchym wie im Pallisadengewebe vor, im ersteren Falle regellos, im letzteren Falle meistens senkrecht zur Blattfläche stehend, Sie unterscheiden sich von den obenerwähnten zu hypodermalen Bündeln vereinigten Faserzellen durch ihren in der Regel stärkeren Durchmesser, sowie verzweigte Gestalt und Vorkommen von Krystallen, Man erkennt ohne Schwierigkeit, dass diese Zellen die Intercellulargänge ausfüllen, und in dieselben hineinwachsend sich verzweigen. Ihre Gestalt ist dementsprechend eine sehr verschiedenartige. Fig. IV. b. zeigt verschiedene Formen der Spicularzellen. Bald sind sie ziemlich einfach mit nur geringen Fortsätzen, bald haben sie eine T- oder H-förmige, bald eine gebogene Gestalt. Was ihre Grosse anbetrifft, so wurden, um dieselbe ungefähr feststellen zu können, Messungen der Zellen in ihrer weitesten Ausdehnung, von einem Ast zum anderen, vorgenommen. Hiernach schwankt ihre Grosse zwischen 590-820 µ. Sie sind deshalb schon mit blossem Auge sichtbar, wenn sie bei sorgfältiger Praeparation aus den umgebenden parenchymatischen Geweben hervorgezogen werden. Auf ihrem Querschnitt, erkennt man, dass in ihrer äussersten Schicht schön ausgebildete Krystalle liegen, welche dem monoklichen System angehören. Dieselben lösen sich bei genügender Einwirkung nach einiger Zeit in Salz- und Salpetersäure, dagegen bleibt Essigsäure wirkungslos. Man kann sich leicht von dieser Thatsache mit Hülfe des Polarisationsapparats überzeugen, welcher bei gekreuzter Stellung der Nicolschen Prismen nach der Behandlung mit diesen Säuren die Stellen, wo die Krystalle sich vorher befanden, dunkel erscheinen lässt. Die Krystalle müssen demnach als Calciumoxalat bezeichnet werden.
Die Resultate der Graf Solms-Laubach'schen Untersuchungen über die Spicularzellen sind in der Einleitung zu diesem Capitel in kurzem wiedergegeben worden.
Ich wandte zur Untersuchung der Spicularzellen folgende Reagentien und Farbstoffe an:
- Für Cellulose.
- Clorzinkjod, welches die bekannte blaue, oder wenn die Cellulose massiv ist, eine tiefbraunrote Reaktion giebt.
- Congorot (in schwach alkoholischer Lösung) welches Cellulose rot, Lignose dagegen gelblich färbt.
- Für Lignin.
- Chlorzinkjod, welches dasselbe gelb färbt.
- Saffranin (in alkoholischer Lösung) mit roter Reaction.
- Fuchsin (in gleichen Teilen Wasser und Glycerin gelöst) mit gleicher Färbung.
- Schwefelsaures Anilin (in Wasser gelöst) Lignin gelb färbend.
- Thallinsulfat (in gleichen Teilen Alkohol und Wasser gelöst) mit orangegelber Reaction auf Lignin.
- Phloroglucin u. Salzsäure (ersteres in Alkohol gelöst). Lignin rot färbend
Thymolsalzsäure, welche ich ebenfalls in Anwendung brachte, blieb ohne wesentliche Einwirkung auf die Wandungen der Spicularzellen, worauf ich im Laufe der Arbeit noch zurückkommen werde,
Ausserdem wurden angewandt als Lösungs- oder Quellungsmittel: Eau de Javelle, Kalilauge, Salzsäure, Essigsäure, Salpetersäure.
Um die Einwirkung dieser Reagentien besser beobachten zu können, wurde ursprünglich nicht mit Schnitten, sondern mit einzelnen freipräparierten Zellen opperiert. Salz- oder Salpetersäure lösen die Krystalle. An ihrer Stelle bleiben in der Wandung der Zellen Löcher zurück. Essigsäure wirkt auf die Krystalle nicht ein. Alle drei Säuren rufen sonst weder bei längerer Einwirkung, bei gewöhnlicher Temperatur noch beim Erwärmen irgend welche Veränderung an der Zelle hervor. Anders verhält sich Eau de Javelle. Hier trat schon nach 4-5stündiger Behandlung der Zellen eine deutliche Quellung der Wandung ein, wie durch vergleichende Messungen festgestellt wurde; auch konnten nach dem Auswaschen der Zelle und Behandlung mit Farbstoffen verschiedenartig gefärbte Schichten an derselben beobachtet werden. Die Zellwandung besteht also aus mehreren chemisch verschiedenen Stoffen, so dass es ausgeschlossen ist, von einer die Wandung bildenden Masse zu sprechen.
Im folgenden sind die Beobachtungen wiedergegeben, welche in verschiedenen Zeiträumen unter Anwendung von Farbstoffen und Reagentien nach oder ohne Behandlung mit Eau de Javelle gemacht wurden:
- Ohne Behandlung mit Eau de Javelle. Mit Chlorzinkjod findet eine langsame nach innen fortschreitende Färbung statt; eine breite äussere Zone wird gelblich, während die übrige Zelle noch ungefärbt bleibt. Erst nach einiger Zeit wird das Innere der Zelle streifig rotbraun und später dunkelviolett, Fuchsin färbt zuerst schwach rötlich und man kann eine Innenzelle und eine breite Aussenzone, welche ungefähr der mit Chlorzinkjod gelb gefärbten entspricht, unterscheiden. Nach vierundzwanzigstündiger Einwirkung dieses Farbstoffes ist die ganze Zelle bis auf einen schmalen Rand, in welchem man viele Krystalle wahrnimmt, dunkel rot gefärbt. Mit Saffranin wird schon nach kürzerer Zeit der Einwirkung die ganze Zelle intensiv rot gefärbt bis auf eine äussere krystallführende Schicht, welche einen schwach bräunlichen Ton annimmt. Mit Congorot tritt nach längerer oder kürzerer Zeit eine Reaction in der Weise ein, dass die ganze Zelle mit Ausnahme der schon erwähnten Aussenschicht rötlich gefärbt wird. Thallinsulfat bewirkt bald eine orangegelbe Färbung der Zelle, nur ein schmaler krystallreicher Aussenrand bleibt farblos. Phloroglucin und Salzsäure färbt die Zelle rot, während der Krystallrand nicht gefärbt wird. Anilinsulfat giebt in kürzerer Zeit eine schön gelbe Reaction; eine schmale Aussenschicht nimmt eine schwach gelbliche Färbung an.
- Nach drei- bis vierstündiger Behandlung mit Eau de Javelle. Mit Chlorzinkjod tritt die unter 1. erwähnte Reaction schneller ein; die gelbe Zone ist also durch die Behandlung mit Eau de Javelle leichter diffundierbar geworden. Als neue Erscheinung ist öfters ein blauer Streifen über der gelben Zone zu bemerken. Ist derselbe nicht vorhanden, so scheinen Krystalle frei auf der Zelle zu liegen. Die übrige Innenschicht der Zellwandung ist in gleicher Weise wie ohne Behandlung mit Eau de Javelle blau gefärbt. Mit Fuchsin wird eine intensive Rotfärbung der ganzen Zelle mit Ausnahme einer dünnen mit Krystallen durchsetzten Aussenschicht auch schon nach kurzer Zeit beobachtet. An einzelnen Stellen ist eine breiige ungefärbte Masse abgequollen, in welcher Krystalle liegen. Mit Saffranin tritt dieselbe Färbung ein; die etwa abgequollene Masse nimmt einen schwach bräunlichen Ton an. Congorot bewirkt rötliche Färbung der ganzen Zelle mit, Ausnahme eines schmalen krystallreichen Aussenrandes, Mit Thallinsulfat tritt bald eine intensive Färbung der Zelle ein; ein ungefärbter au einzelnen Stellen etwas abgequollener Krystallrand ist sichtbar. Anilinsulfat färbt die Zelle ebenfalls gelb, dagegen nimmt der eben erwähnte Krystallrand eine schwach gelbliche Farbe an. Mit Phloroglucin und Salzsäure wird rote Färbung der Zelle hervorgerufen; die Krystallzone bleibt ungefärbt.
- Nach vierundzwanzigstündiger Behandlung mit Eau de Javelle. Mit Chlorzinkjod tritt rasch eine tiefblaue Färbung der ganzen Zelle ein, um welche Krystalle scheinbar frei liegen oder der Zelle noch anhaften, Auch wurden über den Krystallen öfters Stücke einer blau gefärbten Membran bemerkt. Die gelbe Zone, deren bei den vorhergehenden Beobachtungen Erwähnung gethan wurde, war also ganz verschwunden. Wurde eine Zelle, welche mich vier- bis fünfstündiger Behandlung mit Eau de Sarelle und Chlorzinkjod von innen nach aussen eine blau gefärbte, darüber eine gelbe und dann eine dünnere Krystallschicht zeigte, einer weiteren zwanzigstündigen Behandlung mit Eau de Sarelle unterworfen, so ergab sich, dass die gelbe Zone nunmehr ebenfalls blaue Färbung mit Chlorzinkjod angenommen hatte und vergleichende Messungen zeigten, dass in dieser sowohl wie in der Innenzelle keine Quellung stattgefunden hatte. Durch Fuchsin und Saffranin wurde leicht ersichtlich, dass die Krystallschicht zu einer breiten Masse aufgequollen war, welche durch ersteren Farbstoff beinahe farblos blieb, durch letzteren aber eine schwache bräunliche Farbe erhielt, während die übrige Zelle durch beide schön rot gefärbt wurde. Ebenso trat nach Behandlung mit Congorot eine Rotfärbung der Zelle ein, um welche eine beinahe ungefärbte mehr oder weniger abgequollene Masse mit Krystallen lag. Thallinsulfat färbte die Zelle orangegelb, Phloraglucin und Salzsäure rot, Anilinsulfat schön gelb. Die breite abgequollene Masse ist im letzteren Falle schwach gelblich gefärbt.
- Nach achtundvierzigstündiger Behandlung mit Eau de Javelle. Die Zelle ist dunkelblau bei Anwendung von Chlorzinkjod, dunkelrot von Fuchsin und Saffranin. Congorot färbt ebenfalls die ganze Zelle rötlich, Thallinsulfat giebt ihr eine orange-gelbe, Phloroglucin und Salzsäure eine rote, Anilinsulfat eine tiefgelbe Färbung. Die breiige um die Zelle liegende Masse mit Krystallen ist bis auf einige schwache Spuren verschwunden.
Zum genaueren Studium der Spicularzellen wurden nun Quer- und Längsschnitte von Blättern untersucht. Den mikrochemischen Reagentien gegenüber verhielten sich die durchgeschnittenen Spicularzellen folgendermassen:
Fuchsin lässt eine mit einem kleinen Lumen versehene Innenschicht erkennen, um welche eine tiefrote Zone liegt; beide sind geschichtet, während eine über der tiefrot gefärbten Zone mit Krystallen durchsetzte Schicht farblos bleibt, und nicht geschichtet ist, Durch Congorot wird der innerste Teil der Zelle lebhaft rot, die darüberliegende Zone schwächer rot gefärbt. Die Krystallzone bleibt farblos; auf dieser ist aber deutlich eine rot gefärbte Membran sichtbar. Mit Chlorzinkjod tritt eine tiefbraunrote oder blaue Färbung der Innenschicht ein, während die breite gelbe Zone um dieselbe sowohl die mittlere als auch die Krystallschicht umfasst. Über letzterer ist eine blau gefärbte Membran sichtbar, Mit Thallinsulfat oder Phloroglucin und Salzsäure bleiben die Innenschicht wie die Krystallzone ungefärbt. Die mittlere Schicht dagegen wird im ersteren Falle orangegelb, im letzteren schön rot gefärbt. Anilinsulfat lässt die Innenschicht ebenfalls farblos, ebenso wie die Krystallzone nur schwach gelblich gefärbt wird, während die Mittelschicht eine schön gelbe Farbe annimmt.
Aus allen diesen Beobachtungen geht hervor, dass der Bau der Spicularzellen keineswegs ein so einfacher ist, als man bisher annahm. Man muss an denselben folgende Gliederungen unterscheiden, welche aus der Zeichnung Fig. V ersichtlich sind.
Eine "Innenschicht" (a), welche ein kleines Lumen (l) umschliesst. Über dieser liegt ein doppelter Mantel; eine "Mittelschicht" (b) und eine "Krystallschicht" (c). Über beiden befindet sich noch eine besondere "Aussenschicht". Wir wollen in einem Schema die Reactionen der einzelnen Teile kurz noch einmal zusammenfassen:
| Chorzinkjod. | Saffranin. | Fuchsin. | Congorot. | Schwefel-Anilin | Thallinsulf. | Phlorogluc. u. Salzs. | |
|---|---|---|---|---|---|---|---|
| Innenschicht. | blau resp. braunrot. | farblos. | farblos. | rot. | farblos. | farblos. | farblos. |
| Mittelschicht. | gelb. (Nach Behandlung mit Eau de Javelle blau.) | rot. | rot. | rötlich. | tiefgelb. | orangegelb. | rot. |
| Krystallschicht. | gelb. | schwach-bräunlich. | farblos. | farblos. | schwachgelb. | farblos. | farblos. |
| Aussenschicht. | blau. | farblos. | farblos. | rot. | farblos. | farblos. | farblos. |
Conc. Kalilauge führt rasch eine starke Quellung der Krystallschicht herbei. Mit diesem Reagenz behandelte Zellen zerreissen bei leichtem Druck, so dass die Innenschicht, welche deutliche für Cellulose charakteristische gekreuzte Primitivfasern zeigt, sichtbar wird. Auf beiden Seiten dieser Innenschicht liegen dann die Mittelschicht und die stark gequollene Krystallschicht. Kalilauge ungefähr 1:3 verdünnt zeigt im wesentlichen dieselbe Einwirkung wie Eau de Javelle, indem es nach längerer Zeit der Behandlung die Krystallschicht aufquellen, lässt und in der Mittelschicht blaue Reaction mit Chlorzinkjod erzeugt.
Wir müssen also in Anbetracht, dass die angewandten Reagentien und Farbstoffe für die Erkennung von Cellulose und Lignin brauchbare Kriterien abgeben, den Bau der Spicularzellen uns in folgender Weise zusammengesetzt denken:
- Die "Innenschicht" ist geschichtet, neigt deutliche Primitivfaserbildung und besteht aus reiner Cellulose.
- Die "Mittelschicht", ebenfalls geschichtet, besteht aus verholzter Cellulose oder Lignin. Durch Behandlung mit Eau de Javelle und verdünnter Kalilange tritt blaue Reaction der letzteren mit Chlorzinkjod ein.
- Die "Krystallschicht" ist quellbar in conc. Kalilauge. Eine langsame Quellung, welche schliesslich zur Lösung führt, tritt mit Eau de Javelle oder verdünnter Kalilauge ein.
- Die "Aussenschicht" aus reiner Cellulose bestehend.
Es wurde nun der Versuch gemacht, den chemischen Charakter dieser Krystallschicht näher festzustellen. Von der Annahme, dass dieselbe möglicherweise den Proteinstoffen nahe stehe, musste bald Abstand genommen werden, da die hierauf zielenden Reactionen ein negatives Resultat ergaben. Ebenso erwies sich die Vermutung, dass die Krystallschicht eine verschleimte Membran sei, wie sie vielfach im Pflanzenorganismus vorkommt, als irrig, da weder durch längeres Liegen im Wasser noch durch Kochen im letzteren eine Quellung oder Lösung hervorgebracht werden konnte,
Ich begnügte mich schliesslich die allgemeinen chemischen Eigenschaften dieser Krystallschicht festzustellen:
Weder in kaltem noch in heissem Wasser tritt Quellung oder Lösung ein. Conc. Kalilauge bewirkt starke Quellung; eine direkte Lösung konnte nicht beobachtet werden. Verdünnte Kalilauge oder Eau de Sarelle, bewirken unter allmähliger Quellung Lösung. Gegen Säuren, sowohl verdünnte wie concentrirte, verhält sie sich in Wärme und Kälte ziemlich indifferent. Mit Chiorzinkjod tritt eine Gelbfärbung ein. Gegen Farbstoffe ist sie im allgemeinen nicht sehr reactionsfähig, Nur durch schwefelsaures Anilin wird eine schwach gelbliche, durch Saffranin eine schwach bräunliche Färbung hervorgerufen.
Es kann uns nach der Feststellung dieser Krystallschicht nicht wundern, wenn das Calciumoxalat der Spicularzellen sich verhältnismässig schwer in Salz- oder Salpetersäure löst, was vielfach beobachtet wurde. Diese Schicht sowohl wie die sie umgebende Haut sind wohl geeignet, der Lösung der Krystalle einen gewissen Widerstand entgegenzusetzen, wenn auch nicht zu verhindern.
Was nun die aus reiner Cellulose, bestehende Aussenhaut der Spicularzellen betrifft, so ist dieselbe nicht in der Weise aufzufassen, dass sie eine mit den Nachbarzellen gemeinsame Scheide bildet (vergl. pag. 18) sondern sie gehört ganz der Spicularzelle an. Diese Thatsache kann bei geeigneter Färbung mit Congorot deutlich beobachtet werden. Auch geht, es aus dem Umstände hervor, dass eine freipräparierte Zelle immer scharf durch diese Membran begrenzt ist ohne jegliche Anhängsel, welche doch im anderen Falle notwendiger Weise als Fetzen der Membranen der Nachbarzellen mit herausgerissen werden müssten.
Die Untersuchung der Spicularzellen im Stamm der Welwitschia mirabilis ergab, dass dieselben den gleichen Bau wie die des Blattes besitzen. Für unterscheiden sie sich von jenen durch ihre wenig verzweigte Gestalt, sowie durch ihre grösseren Dimensionen (Fig. V. a). Sie übertreffen die des Blattes um das zwei- bis dreieinhalbfache an Grösse und erreichen eine Länge von 1,55-1,85 mm.
Cap. II.
Das Calciumoxalat in den Idioblasten der Coniferen.
Die der Welwitschia mirabilis nahestehenden Coniferen besitzen in Stamm und Blatt der Spicularzellen dieser ersteren sehr ähnliche Krystallidioblasten. Dieses gemeinsame Vorkommen eines so charakteristischen Gewebestücks vermehrt. die vielfach anatomischen Ähnlichkeiten dieser habituell so verschiedenen Gewächse, welche auch ich bei der Untersuchung der Blätter von Coniferen zu beobachten Gelegenheit hatte. Besonders auffallend ist der ähnliche Bau des Spaltöffnungsapparats und das Vorkommen der starken Sclerenchymfasern im Hypoderm sowohl als auch als Schutzscheide der Gefässbündel, welche Verhältnisse bei der Anatomie der Welwitschia eingehend besprochen worden sind. Von den von mir untersuchten Krystallidioblasten der Gattungen Araucaria, Sciadopitys und Dammara haben nur die ersteren von Winkler [35] eine eingehendere Besprechung erfahren. Derselbe giebt, in seiner Abhandlung an, dass schon bei der Differenzierung der Gewebe die Zellen mit unzählig kleinen Pünktchen bedeckt seien, unter denen zuweilen auch schon ein Krystall sichtbar wäre. Die Einschliessung der Krystalle wird nach ihm durch Überwallung der Membran hervorgerufen, da sich Übergänge von anscheinend lose aufsitzenden bis zu solchen Krystallen finden, welche vollständig in der Membran sitzen.
Die Araucarien führen im Blatt und in der Stammrinde Krystallidioblasten. Im ersteren Falle sind sie spärlich vertreten und deshalb zu einer Untersuchung nicht recht geeignet.
Häufig dagegen sind sie in der Stammrinde und zwar in zweierlei Form. Im Weichbast finden wir unverzweigte, spindelförmige bis 4,4 mm. lang werdende Krystallidioblasten mit deutlicher Schichtung und Tüpfelkanälen. Das Lumen kann verschieden ausgebildet sein; doch ist es meistens klein und wird durch die Verdickungen vollständig verdrängt. Man kann im letzteren Falle eigentümlicher Weise oft eine breite Aussen- und eine Innenzone, welche scharf von einander begrenzt sind, unterscheiden. Zweitens beobachtet man Krystallidioblasten im Rindenparenchym. Sie sind von annähernd isodiametrischer Gestalt, mit kurzen Fortsätzen versehen. Sie zeigen deutliche Schichtung und sind mit Tüpfelkanälen versehen. Ihr Lumen war in den beobachteten Fällen noch verhältnismässig gross. Sie werden 272-300 µ. lang. Untersucht wurden von mir die beiden Spec.: Araucaria brasiliana Lamb. und Araucaria imbricata Pav. Die im Mesophyll des Blattes von Sciadopitys verticillata Letz. zahlreich vorkommenden Krystallidioblasten zeigen geschichtete Verdickungen, aber keine Tüpfelkanäle. Sie erreichen eine Lunge von 372-390 µ.
Agathis Dammara Rich. besitzt ebenfalls im Mesophyll des Blattes Krystallidioblasten. Diese sind den Spicularzellen des Welwitschiablattes ähnlich, aber insofern abweichend in ihrer äusseren Form gebaut, als man bei ihnen einen rundlichen oder länglichen Hauptteil und von diesen ausgehende spitzenartig unregelmässig nach allen Richtungen gehende Fortsätze unterscheiden kann. Sie zeigen Schichtung und Tüpfelkanäle. Ihre Verdickungen sind bei alten Blättern bis fast zum vollständigen Schwinden des Lumens vorgeschritten. Sie werden durchschnittlich 225-240 µ. lang. Die Krystalle des Calciumoxalats sind in allen diesen untersuchten Fällen nicht der Membran, sondern einer eigenen Krystallschicht eingelagert, welcher dieselben chemischen Eigenschaften zukommen, wie der bei den Blattspicularcellen von Welwitschia beobachteten. Eine besondere Aussenhaut von Cellulose um
diese Schicht konnte wie bei jenen nicht beobachtet werden. Die Verdickungen der Zellen bestehen mit einer Ausnahme aussen aus verholzter, innen aus reiner Cellulose, wie mit allen den obenerwähnten (pag. 25) Reagentien und Farbstoffen nachgewiesen wurde, Nur die unverzweigten, spindelförmigen Idioblasten im Weichbast der Araucarien bestehen in ihrer ganzen Masse aus verholzter Cellulose. Letztere giebt in allen beobachteten Fällen nach vierundzwanzigstündiger Behandlung mit Eau de Javelle oder verdünnter Kalilauge blaue Cellulosereaction mit Chlorzinkjod,
Ich hatte nun auch Gelegenheit die Krystallidioblasten im Blatt von Agathis Dammara Rich. entwicklungsgeschichtlich zu verfolgen. Im hiesigen botanischen Garten war ein, prächtiges Exemplar dieses Baumes vorhanden, welches mir zur Untersuchung gütigst überlassen wurde, so dass ich in der Lage war, Blätter in allen Entwickelungsstadien zu untersuchen.
In sehr jungen Blättern, welche die Dicke von 280 µ. noch nicht überschritten haben, sind die Gewebe zum Teil noch nicht fertig ausgebildet. Das spätere Pallisadengewebe besteht noch aus isodiametrischen Zellen, während das den grössten Teil des Blattes einnehmende Schwammparenchym ans rundlichen Zellen besteht. In letzteren sind noch keine Intercellulargänge sichtbar. Gefässbündel und Harzgänge sind schon fertig ausgebildet. Krystallidioblasten bemerkt man noch nicht, doch fallen kleine meist viereckige Zellen auf, welche sich mir durch ihre geringere Grösse von den übrigen Zellen unterscheiden und sich möglicherweise zu Idioblasten entwickeln, was jedoch eine besondere Untersuchung feststellen müsste. Betrachtet man nun ein älteres Blatt von der durchschnittlichen Breite von 280-320 µ., so zeigt sich, dass das noch unfertige Pallisadengewebe in beginnender Differenzirung und Streckung sich befindet. Ton den Zellen, welche um die nun auch sichtbar werdenden Intercellulargänge herumliegen, treiben einige Fortsätze, in diese hinein. Solche Zellen schwanken
in ihrer Länge zwischen 54-140 µ. Im Übrigen unterscheiden sie sich in keiner Weise von den übrigen Zellen, des sie umgebenden Gewebes; sie besitzen Protoplasma und ihre Wandung besteht aus reiner Cellulose. Die Beobachtung eines noch älteren Blattes in der Dicke von 320-300 µ., ergiebt, dass das Wachstum dieser Zellen bedeutend fortgeschritten ist, denn sie haben jetzt durchschnittlich eine Länge von 215 bis 245 µ. erreicht. Die Membran reagiert nur schwach auf Cellulose, giebt dagegen mit Thymolsalzsäure eine blaugrüne Färbung, wodurch die beginnende Verholzung durch Gegenwart des durch letzteres Reagenz angezeigten Coniferins festgestellt wird. Ein Protoplasma konnte auf Schnitten nicht beobachtet werden. Ebenso zeigten die unter Zugabe von etwas Wasser frei präparierten Zellen kein Protoplasma und erschienen sogar vielfach mit Luft erfüllt. Erst bei Präperation der Zellen unter erstarrender Glyceringelatine, wo also der Zutritt von Luft und Wasser ausgeschlossen war, konnte in diesem Stadium der Zellen und auch in späteren ein Protoplasma beobachtet werden. Dieses Verhalten des letzteren kann nur in der Weise erklärt werden, dass es sich zu dieser Zeit und auch späterhin in anormalem Zustande befindet, in welchem es sehr leicht, aus der Zelle ausfliesst. Ein Blatt in der Dicke von 380-440 µ. lässt erkennen, dass die Idioblasten an Grösse nicht zugenommen haben, aber ihre verholzte Wandung oft schon Ansätze zu Tüpfelkanäle und Krystalle zeigte. Eine weitere Untersuchung eines Blattes von -600 µ. Dicke, welches als ausgewachsen zu betrachten ist, zeigt, dass ein nachträgliches Wachstum der Idioblasten nicht stattfindet. Die Wandung derselben aber ist stark verdickt mit Schichtung und Tüpfelkanälen versehen. Ihrer chemischen Natur nach besteht der Innenrand aus reiner Cellulose, der Aussenrand der Verdickungen aus verholzter Cellulose. Die letztere giebt auch schon bei Beginn der Verdickungen und der Krystallbildung, sowie späterhin keine Reaction mit Thymolsalzsäure, dagegen eine schöne orangegelbe mit Thallinsulfat, wodurch Vanillin im Holz angezeigt ist. Auf der Zelle liegen in der Krystallschicht zahlreiche Calciumoxalatkrystalle.
Die Entwicklungsgeschichte der Krystallidioblasten im Blatt von Agathis Dammara verläuft also in folgenden Stadien:
Zu der Zeit, in welcher das Pallisadengewebe noch nicht ausgebildet ist, unterscheiden sich die zu Idioblasten werdenden Zellen nicht besonders von den sie umgebenden des Schwammparenchyms. Erst wenn das Pallisadengewebe sich zu strecken anfängt, beginnen Zellen in die sich nun ebenfalls bildenden Intercellulargänge hineinzuwachsen, erreichen rasch eine ziemliche Grösse und stellen dieses Wachstum ungefähr zu der Zeit ein, in welcher auch das Pallisadengewebe sich ausgebildet hat. Zu dieser Zeit und auch später ist das Protoplasma schwer nachweisbar. Kurz darauf beginnt die Verholzung der Membran, welche kurze Zeit Coniferinreaction und dann Vanillinreaction zeigt, sowie die Krystallbildung. Die Verdickungen schreiten von aussen nach innen vor; damit steht im Zusammenhang die Krystallbildung, so dass, je stärker die Verdickungen, um so reichlicher die Krystallausscheidung vor sich geht.
Cap. III.
Das Calciumoxalat in den Idioblasten der Nymphaeaceen.
Die Krystallidioblasten der Nymphaceen ragen in die grossen dieser Familie eigenen Intercellulargänge hinein und sind von sehr verzweigter Gestalt. Kohl [36] führt gegenüber der Behauptung von Molich [37] aus, dass die Krystalle des Calciumoxalats hier der Membran eingelagert und nicht aufgelagert seien. Es müssten im letzteren Falle bei Einwirkung von Salzsäure die Stellen, an welchen die Krystalle liegen, nicht hervorgewölbt bleiben, was jedoch stattfindet. Er ist wie Schenk [38] der Ansicht, dass das Calciumoxalat von dem Protoplasma an der Innenseite der primären Wand abgeschieden und von den später gebildeten Verdickungsschichten nach aussen geschoben wird.
Untersucht wurden von mir die Krystallidioblasten von Nymphaea alba L. und Nuphar luteum L. Die meistens nur schwach verdickten Zellen zeigen deutliche Schichtung. Die Wandung derselben ist an denjenigen Stellen, mit welchen sie an die, Nachbarzellen stösst, eigentümlich gelappt, welche Erscheinung jedenfalls durch ein ungleiches Wachstum bedingt ist. Die Idioblasten von Nymphaea alba werden 400-450 µ. lang, diejenigen von Nuphar luteum erreichen eine Grösse von 400-425 µ.
Auch hier sind die Krystalle in eine besondere nach aussen liegende, wie bei den vorher beschriebenen Idioblasten, Schicht eingebettet. Der äussere Teil der Zellwand ist verholzt, wie mit den oben erwähnten Reagentien und Farbstoffen (pag. 25) nachgewiesen wurde, und nimmt nach eintägiger Behandlung mit Eau de Javelle durch Chlorzinkjod blaue Reaction an. Der Innenrand der Zellwandung besteht aus Cellulose.
Nach diesen Ausführungen ist man wohl berechtigt, die als "Sternhaare" bezeichneten verzweigten Zellen der Nymphaceen zu den Krystallidioblasten zu rechnen, da sie sich in nichts wesentlichem von jenen unterscheiden. Nur können wegen ihrer nur einjährigen Ausdauer die Verdickungen der Idioblasten nicht so weit vorschreiten, wie in den anderen beobachteten Fällen.
Zur Physiologie der Krystallidioblasten.
Ziehen wir nun allgemeine Schlüsse aus den erwähnten Beobachtungen und der entwickelungsgeschichtlichen Untersuchung der Krystallidioblasten, so ergiebt sich folgendes: Ihre Gestalt ist im wesentlichen abhängig von den Intercellulargängen, in denen sie hinwachsen. Man findet deshalb in der Rinde lange spindelförmige, wenig oder garnicht verzweigte, dagegen im lockeren Schwammgewebe der Blätter vielfach verästelte Idioblasten. Sie sind ungefähr zu der Zeit ausgewachsen, in welcher alle anderen Gewebe differenzirt sind, zeigen aber in diesem Stadium noch keinerlei Verdickung, Erst später beginnt, die Verholzung und das Verdicken der Zellwand, welches unter Umständen zum fast vollständigen. Schwinden des Lumens führen kann.
Es entsteht nun die Frage, ob das Protoplasma der Idioblasten im Stande ist, solche starke Verdickungen zu bilden? Mir scheint, dass hier wandernde Kohlenhydrate, welche im Protoplasma aufgespeichert werden, das Material zu den Verdickungen bilden, Und zwar gelangen diese Kohlenhydrate in Form eines Glycosekalks in die Zellen nach Kohl's [39] Ansicht. Diese vielfach angenommene und von ihn selbst, ausgesprochene Wanderung der Kohlenhydrate in Form einer Kalkverbindung glaubt Schimper [40] wie in der Einleitung angegeben in allerneuster Zeit nicht annehmen zu dürfen. Nach Kohl wird an allen denjenigen Stellen, wo ein Kohlenhydrate aus seiner Kalkverbindung frei wird, gleichzeitig Kalk disponibel werden, welcher sich mit der in den Pflanzen gebildeten Oxalsäure zu oxalsaurem Kalk verbindet. Zu Folge dessen muss sich in der Nahe grösserer Cellulosemassen immer Calciumoxalat finden. Dies ist in der That der Fall, wie Kohl vielfach nachgewiesen hat. Auch meine Beobachtungen stimmen mit dieser Ansicht gut überein. Besonders charakteristisch ist, ein neben vielen anderen von Kohl angeführter Fall betreffend die Schalen vieler Monocotyledonenzwiebeln. Jede Epidermiszelle enthält hier einen grossen Calciumoxalat-Krystall. Je mehr nun Cellulose in den Wänden des hypodermalen Gewebes gebildet wird, um so zahlreicher sind die Krystalle des Oxalats. Dieser Fall passt genau zu den Verhältnissen, wie sie bei den Krystallidioblasten beobachtet wurden, bei denen ebenfalls eine fortschreitende Verdickung der Zellwandung durch die Zunahme des Calciumoxalats bedingt ist. Ferner sind als bemerkenswert zu erwähnen die Beobachtungen Garcin's [41], welcher in den dreischichtigen Steinkernen von Crataegus oxyacantha und Svmphoricarpus racemos. in den stark verdickten Zellen der mittleren Sclerenchymschicht je einen Krystall von Calciumoxalat fand. Auch Mattirolo [42] hat bei der Entwickelung von Samenknospenhüllen der Tilia-Arten beobachtet, dass Zellen aufhören sich zu teilen, ihr Inhalt unter gleichzeitiger Verdickung der Zellwand schwindet und dabei zugleich reichlich oxalsaurer Kalk im Inneren der Gewebe sich bildet. Wir sind auch mithin berechtigt, anzunehmen, dass die Verdickungen der Krystallidioblasten durch die wandernden Kohlenhydrate gebildet werden. Da die Idioblasten sich erst anfangen zu verdicken, wenn die sie führenden und benachbarten Gewebe vollständig, oder doch nahezu vollständig ausgebildet sind, so scheinen die die Verdickungen bildenden einwandernden Kohlenhydrate keine Verwendung für den Aufbau der Gewebe der Pflanze mehr zu haben. In welcher Weise nun die Glycose vom Kalk getrennt und in Cellulose verwandelt wird, darüber haben wir bis jetzt wenig Anhaltspunkte. Möglicher Weise findet dieser letztere Prozess nach Ebermayer [43] durch Condensation zweier Moleküle Zucker unter Wasserabspaltung statt:
2 C6H12 - H2O = C12H20O10
Auch hat Durin, wie Ebermayer anführt, gezeigt, dass in der Zuckerrübe ein der Diustase nahestehendes Ferment vorhanden ist, welches in diesem Falle Rohrzucker in Cellulose verwandelt, indem sich ersterer in Cellulose und Laevulose spaltet, Man kann nun im allgemeinen annehmen, dass bei der Umwandlung der Kohlenhydrate in Cellulose ein im Protoplasma enthaltenes Ferment wirksam ist. Wir erinnern daran, dass, wie oben angegeben, zur Zeit der Ausscheidung der Verdickungen in den Idioblasten ein Plasma zwar vorhanden, aber schwer nachweisbar ist. Was nun die mit den zunehmenden Verdickungen vermehrte Bildung des Calciumoxalats betrifft, so entspricht dasselbe dem von Kohl [44] tertiär genannten, welches in der Nähe grösserer Cellulosemassen vorkommt. Die Oxalsäure denke ich mir durch Oxydation der verholzten Cellulose der Idioblasten entstanden, welcher Prozess makrochemisch leicht ausgeführt werden kann, Es ist eine bekannte Thatsache, dass die fabrikmässige Darstellung der Oxalsäure in einem solchen Oxydationsprozess aus Holz besteht, indem Sägespäne mit einem Gemisch von Kali- und Natronhydrat erhitzt werden, wobei oxalsaures Alkali resultiert. Die erstgebildeten Verdickungen und mithin äussersten Schichten der Idioblasten sind, was bemerkenswert ist, in allen beobachteten Fällen verholzt. Bis jetzt ist es nicht möglich gewesen, den Holzstoff chemisch scharf zu definiren. Die Ansichten der einzelnen Forscher [45] differieren recht stark. Nur soviel ist sicher, dass wir den Holzstoff nicht als eine einzelne, sondern als ein Gemenge chemischer Verbindungen aufzufassen haben. Als Grundsubstanz ist Cellulose zu betrachten, welcher die xylogenen chemischen Verbindungen, die wir kurz als "Lignose" bezeichnen wollen, molekular eingelagert sind. In neuerer Zeit ist man der Kenntnis dieser xylogenen Verbindungen nähergetreten, indem Singer [46] feststellte, dass das Coniferin und das aus ihm hervorgehende Vanillin immer im Holz enthalten seien und durch das Auftreten dieser beiden in der Cellulosemembran die Holzbildung bedingt sei. Das Coniferin ist ein Glycosid und war schon längere Zeit, als im Cambialgewebe von Coniferen vorkommend bekannt. Tiemann [47], welcher in einer Reihe von Arbeiten das Coniferin in chemischer Beziehung untersucht hat, fand, dass dasselbe bei Einwirkung von verdünnten Säuren, insbesondere aber von ungeformten Fermenten, z. B. dem Emulsin in Coniferylalcohol und Traubenzucker gespalten wird. Bei Oxydation des ersteren entsteht das Vanillin oder Methylprotocatechualdehyd, derjenige Stoff, welcher der Vanille den angenehmen Geruch verleiht. Hegeler [48] sucht den Prozess der Bildung des Coniferins durch eine chemische Formel auszudrücken und fuhrt den späteren Zerfall des Coniferins in Coniferylalcohol und Traubenzucker auf die fermentartige Beihülfe "ungeformter Eiweisskörper protoplasmatischen Ursprungs zurück". Es gelang ihm auch besonders durch vergleichende Versuche mit Coniferin und Vanillin nachzuweisen, dass Thallinsulfat nur mit letzterem, Thymolsalzsäure nur mit ersterem reagiere, während alle anderen Verholzung anzeigenden Reagentien mit beiden Körpern Farbenreactionen hervorbrächten. Es lässt sich damit leicht unterscheiden, ob ein Holz Coniferin oder aus diesem durch Oxydation hervorgegangenes Vanillin enthält. Hegeler konnte mithin auf Querschnitten verschiedener Stammhöhen derselben Pflanze feststellen, dass die dem Vegetationspunkt zunächst liegenden Holzgewebe viel Coniferin, dagegen wenig oder gar kein Vanillin enthalten, während bei weiter vom Vegetationspunkt gelegenen Holzgeweben das umgekehrte der Fall ist. Es hat also im letzteren Falle schon eine Oxydation zu Vanillin stattgefunden. Die Verholzung der Krystallidioblasten zeigt, wie in den vorhergehenden Untersuchungen festgestellt wurde, starke Vanillinreaction mit Thallinsulfat. Die Thymolsalzsäurereaction auf Coniferin tritt, wie die entwickelungsgeschichtliche Untersuchung der Blattkrystallidioblasten von Agathis Dammara zeigt, nur ein in dem Stadium der Zelle, in welcher die Verholzung beginnt. Sobald die Verdickung der Zellwand, sowie die Krystallbildung anfängt, waltet bereits die Vanillinreaction vor.
Die von verschiedenen ausgeführte Elementaranalyse des Holzstoffs hat nicht allzu differiernde Resultate ergeben, von denen hier das eine angeführt ist:
C = 55,55 % }
H = 5,83 % } Schulze.
O = 38,62 % }
Man hat sogar versucht eine Formel für den Holzstoff aufzustellen. Entweder wird C18H21O10 oder C10H24O10 angenommen. Diese Formel hat natürlich nur insofern Wert, als sie uns das relative Verhältnis der einzelnen Elemente angiebt. Mit der Ausführung dieser Elementaranalyse ist aber sicher nachgewiesen, dass der Holzstoff relativ kohlenstoffreicher, aber sauerstoffärmer als Cellulose ist. Die procentuale Zusammensetzung der letzteren ist folgende:
C = 44,44 %
H = 6,17 %
O = 49,39 %
Nach Ebermayer [49] ist der Holzstoff als ein Umwandlungsprodukt der Cellulosemoleküle zu betrachten, welche möglicherweise auf einer durch Atmung bewirkten Oxydation der Cellulose beruht, wobei unter Wasserabspaltung eine kohlenstoffreichere Verbindung zurückbleibt;
4 C6H10O5 + 10 O = C18H24O10 + 6 CO2 + H2O.
Die Verholzung der Zellwand bei den Idioblasten schreitet von aussen nach innen vor, da die äusseren Schichten verholzt, die inneren dagegen im allgemeinen aus Cellulose bestehen. Ich nehme nun an, dass diese verholzte Zellwand, welche schon vorher einen Oxydationsprozess in der Umwandlung des Coniferins in Vanillin in sich vollzogen hat, in ihren äussersten Schichten eine weitere Oxydation erfährt zu der Zeit, in welcher die von den Zellen aufgenommenen Kohlenhydrate in Form von Cellulose auf der Innenseite der Zellwand abgeschieden werden. Dieser Oxydationsprozess verläuft analog demjenigen, welcher sich bei der technischen Verarbeitung von Holz zu Oxalsäure abspielt: Unter alkalischer Einwirkung von Kalk und Hinzutritt von Sauerstoff bildet sich aus den äussersten Schichten der verholzten Cellulose durch Vermittelung der Krystallschicht, welcher besondere chemische Eigenschaften zukommen, Calciumoxalat.
Es findet demnach eine allmählige Überführung der Cellulose in Oxalsäure durch Oxydation statt, wobei zuerst eine kohlenstoffreichere Verbindung resultiert, welche dann unter Einwirkung von Kalk und Sauerstoff das Endprodukt der Oxydation der Kohlenhydrate, die Oxalsäure, liefert.
Es entsteht nun die Frage, ob in der Pflanze solche Oxydationen, welche in der Makrochemie doch kräftiger Oxydatoren und höherer Temperaturen bedürfen, durch den durch die Atmung zugeführten Sauerstoff bewirkt werden können, Reinke [50] hebt in seiner Abhandlung hervor, "dass zwar eine reine Traubenzuckerlösung bei niedriger Temperatur durch den indifferenten, molekularen Sauerstoff nicht oxydiert wird, während nach unserer Auffassung eine solche im lebenden Protoplasma verteilte Lösung unter Absorption des Sauerstoffs eine totale Verbrennung erfährt". Ferner gelangt er im Laufe derselben Arbeit zu Resultaten, welche er in folgenden Sätzen wiedergiebt:
- "In jeder lebensfähigen Zelle werden Autooxydatoren d. h. Substanzen gebildet, welche bei niedriger Temperatur unter Aufnahme von molekularem Sauerstoff aus dem Medium durch Wasserzersetzung oxydiren."
- "Bei der Oxydation dieser Autooxydatoren entsteht aus dem dabei mitwirkenden Sauerstoff Wasserstoffsuperoxyd."
- "Dieses Wasserstoffsuperoxyd vermag unter Einwirkung von Diastase, wahrscheinlich auch anderer Fermente, Oxydationen von ähnlicher Energie auszuführen wie der atomistische Sauerstoff."
Nach diesen Untersuchungen können wir annehmen, dass nicht der passive, bei der Atmung aufgenommene Sauerstoff, sondern der vom Protoplasma activ gemachte Sauerstoff im Stande ist, solche kräftige Oxydation unter alkalischer Mitwirkung des Kalks auszuführen.
Sucht man die Bedeutung der Krystallidioblasten für die Biologie der Pflanze auf, so ist das Vorkommen derselben als zweckmäßig zu betrachten, als in ihnen die für die Pflanze nicht mehr nutzbaren Kohlenhydrate als Cellulose aufgespeichert werden. Auf ihrer Wanderung werden diese Kohlenhydrate in gelöster Form der Kalkverbindung in den Krystallidioblasten angehäuft. Der bei der Ausscheidung der Kohlenhydrate in Form von Cellulose freiwerdende Kalk wird durch die sich gleichzeitig bildende Oxalsäure gebunden. Die Bedeutung der stets isolirten Krystallidioblasten für das mechanische System kommt überhaupt nicht in Betracht. Denn zur Aussteifung der Gewebe sind bei den Pflanzen, welche Krystallidioblasten führen, meist hinreichend andere sclerenchymatische Elemente vorhanden, z. B. im Blatt der Welwitschia mirabilis die starken hypodermalen Sclerenchymfaserbündel, welche für sich allein befähigt sind, dem Blatt die nötige Festigkeit zu verleihen.
Zusammenfassung der Ergebnisse.
Fassen wir nun am Schluss die aus den Untersuchungen hervorgehenden Resultate noch einmal kurz zusammen, so wurde folgendes festgestellt:
1) Die Krystallidioblasten von Welwitschia mirabilis Hook., der Coniferen (Agathis Dammara Rich., Araucaria imbricata Pav., Araucaria brasiliana Lamb., Sciadopitys verticillata Letz.) und der Nymphaeaceen (Nymphaea alba L. Nuphar luteum L.) zeigen im allgemeinen einen übereinstimmenden Bau. Sie sind mehr oder weniger verzweigt. Ihre Verdickungen bestehen aus chemisch verschiedenen Schichten, deren innere ans reiner Cellulose, die äussere aus verholzter Cellulose besteht, welche letztere nach vierundzwanzigstündiger Behandlung mit Eau de Javelle oder verdünnter Kalilauge blaue Reaction mit Chlorzinkjod giebt, Auch erhält man mit Thallinsulfat eine orangegelbe Reaction der verholzten Cellulose, wodurch die Gegenwart von Vanillin angezeigt ist, während Thymolsalzsäure, Coniferin anzeigend, wirkungslos auf dieselbe bleibt. Die spindelförmigen Krystallidioblasten im Weichbast der Araucarien bestehen nur aus verholzter Cellulose, Auf der Zelle liegen in einer besonderen chemisch nicht scharf zu charakterisirenden Schicht die Calciumoxalatkrystalle. Dieselbe ist quellbar in conc. Kalilauge, ausserdem auch noch löslich in Eau de Javelle oder verdünnter Kalilauge, Bei den Idioblasten von Welwitschia mirabilis befindet sich um die Krystallschicht noch eine aus reiner Cellulose bestehende Aussenhaut. Wie die entwickelungsgeschichtliche Untersuchung im Blatt von Agathis Dammara zeigt, entstehen die Krystallidioblasten aus Zellen des Parenchyms, welche sich durch nichts wesentliches von den sie umgebenden unterscheiden. Sie beginnen zu einer Zeit, in welcher auch andere Gewebe noch nicht vollkommen differenziert sind, in die Intercellulargänge auszuwachsen. Sie erreichen ihre definitive Grösse ungefähr zu der Zeit, in welcher auch alle anderen Gewebe ausgebildet sind, ohne dabei irgend wie die Zellwand zu verdicken. Nach dieser Zeit ist ein Protoplasma zwar vorhanden, aber schwer nachweisbar. Darauf beginnt die Zellwand zu verholzen, wobei sie zuerst kurze Zeit Coniferinreaction mit Thymolsalzsäure giebt, um allerdings nach kurzem die Vanillinreaction mit Thallinsulfat zu zeigen. Es findet dann eine fortschreitende Verdickung nach innen zu statt. Je stärker die Verdickungen werden, um so mehr Calciumoxalat wird in der Krystallschicht ausgeschieden. Was die Physiologie der Krystallidioblasten betrifft, so ist anzunehmen, dass die Cellulose der Verdickungen aus den von der Zelle osmotisch aufgenommenen flüssigen Kohlenhydrat-Kalkverbindungen herstammt. Die Trennung des Kohlenhydrats vom Kalk und die Abscheidung des ersteren in Form von Cellulose erfolgt wahrscheinlich durch fermentartige Wirkung des Protoplasma. Der frei werdende Kalk wird durch Oxalsäure zu Calciumoxalat gebunden. Die Oxalsäure ist als ein Oxydationsprodukt der äussersten verholzten Zellschichten der Idioblasten, welche bereits einen Oxydationsprozess des Coniferins zu Vanillin in sich vollzogen haben, zu betrachten unter alkalischer Mitwirkung von Kalk und gleichzeitigem Hinzutritt von Sauerstoff, welchem durch das Protoplasma eine höhere Oxydationskraft erteilt wird. Die Ausscheidung erfolgt, wie erwähnt, in der Krystallschicht, welche ihrerseits als ein intermediäres Produkt zwischen Cellulose und Oxalsäure zu betrachten ist. Die biologische Bedeutung der Krystallidioblasten ist in erster Linie darin zu suchen, dass überflüssige Kohlenhydrate in Form von Cellulose in ihnen ausgeschieden wird. In zweiter Linie unterstützen sie vielleicht das mechanische System, wofür ihr isolirtes Vorkommen allerdings nicht spricht.
2) Zur Anatomie des Blattes von Welwitschia mirabilis Hook. haben meine Untersuchungen folgendes ergeben:
Die stark gewellte Epidermis besitzt längliche stark verdickte Zellen. Zwischen diesen und der cuticula liegt eine körnige, aus oxalsaurem Kalk bestehende Schicht, welche sich durch ihr Verhalten gegen Farbstoff als chemisch verschieden von der krystallfreien cuticula und der Wandung der Epidermiszellen erweist. Sie ist deshalb weder der ersteren noch der letzteren zuzurechnen und wird am besten "Krystallogenschicht" genannt. Die Epidermis besitzt an den Spaltöffnungen eine doppelte Einsenkung, so dass ein Vor- und Hinterhof gebildet wird. Die Schliesszellen der Spaltöffnung sind auf ihrer Vorder- und Hinterseite mit einer Einschnürung versehen. Die Verdickungen, welche in der Mitte der Schliesszellen am stärksten, nach unten und oben zu schwächer werden, liegen ebenfalls an der Vorder- und Hinterseite der Schliesszellen. Das Öffnen des Spaltöffnungsapparates erfolgt durch ein passives Auseinanderweichen der Verdickungen auf der Vorder- und Hinterseite und eine dadurch bewirkte Annäherung der schwach oder garnicht verdickten Bauch- und Rückenseite (vergl. pag. 19). Abweichend vom allgemeinen Bau der Blätter liegt auf der Ober- und Unterseite des Welwitschiablattes Pallisadengewebe. In den Intercellularräumen desselben beobachtet man in jungen Blättern grosse Mengen kleiner Calciumoxalatkrystalle. Den älteren Blättern fehlt diese Erscheinung. Die Intercellularräume des aus rundlichen Zellen bestehenden Blattparenchym sind ebenfalls ausgefüllt mit körnigem oxalsaurem Kalk, welcher dem Gewebe ein scheinbar collenchymatisches Aussehen verleiht. In diesem Teil des Blattes liegen auch die durch Anastomosen verbundenen Gefässbündel. Sie sind collateral und umgeben von einem aus genetzten und getüpfelten Zellen bestehenden "Transfusionsgewebe" oder "Tracheidensaum", welcher nicht zum Gefässbündel gehört. Das eigentliche Gefässbündel besitzt zwei halbmondförmige Sclerenchymscheiden, das der Unterseite des Blattes zugewendete "Phloëm" besteht aus engmaschigem meist zusammengedrückten Gewebe, welches keine Formbestimmung seiner Elemente zulässt. Das der Oberseite zugewendete "Xylem" besteht aus dickwandigen lückenlos verbundenen Zellen, deren Wandung besonders in der Nähe der Sclerenchymscheide stark gequollen ist. Zwischen diesem Gewebe befinden sich fast in der Mitte des ganzen Gefässbündels die Tracheen; einige kommen auch versprengt in der Nähe der Sclerenchymscheide vor. Die dem Phloëm zunächst liegenden Tracheen sind ächte Netz- und Tüpfelgefässe, die darauf folgenden dagegen Ring- und Spiraltracheiden. Die Anastomosen sind ähnlich gebaut wie die Gefässbündel. Doch ist das gequollene Gewebe des Xylems schwach entwickelt und der Tracheidensaum bis auf einige wenige genetzte und getüpfelte Zellen reducirt. Die Sclerenchymscheiden fehlen vollständig.
Zeichnungen
Figur I
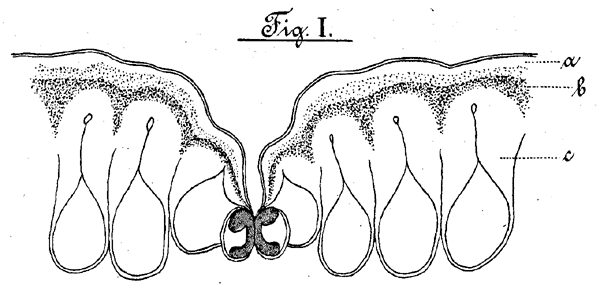
Querschnitt durch die Epidermis des Welwitschiablattes (vergr. 1:585).
a. cuticula, b. Krystallogenschicht, c. Epidermiszellen.
Figur II
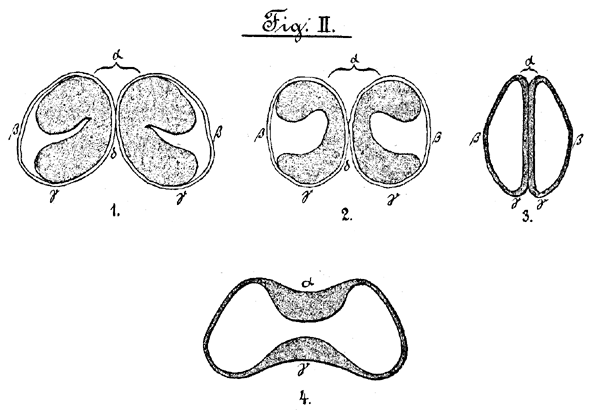
Schliesszellen des Spaltöffnungsapparates des Welwitschiablattes
(vergr. 1:790).
1.-8. Querschnitte bei verschiedenen Einstellungen. 4. Längsschnitt.
α Vorderseite, β Rückenseite, γ Hinterseite,
δ Bauchseite.
Figur III
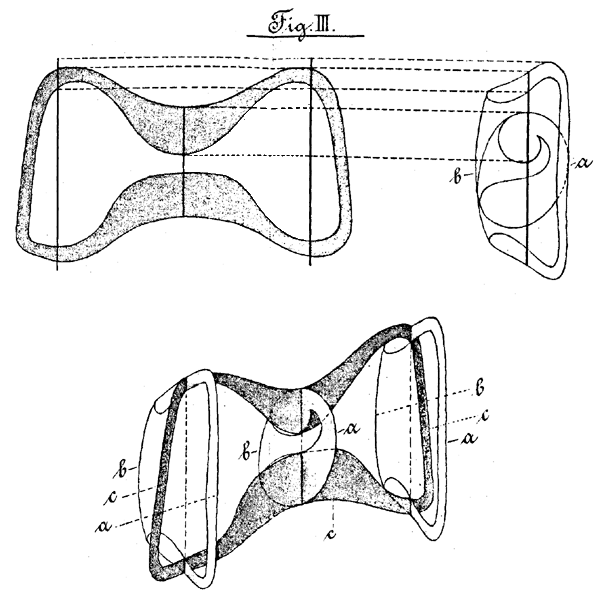
Schematische Darstellung der Verdickungen der Epidermiszellen.
a. Bauchseite, b. Rückenseite, c. Vorder- und Hinterseite.
Figur IV
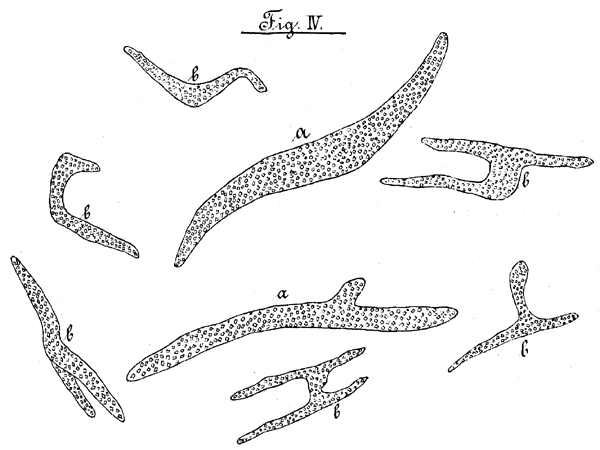
Spicularzellen der Welwitschia (vergr. 1:58).
a. aus dem Stamm, b. aus dem Blatt.
Figur V
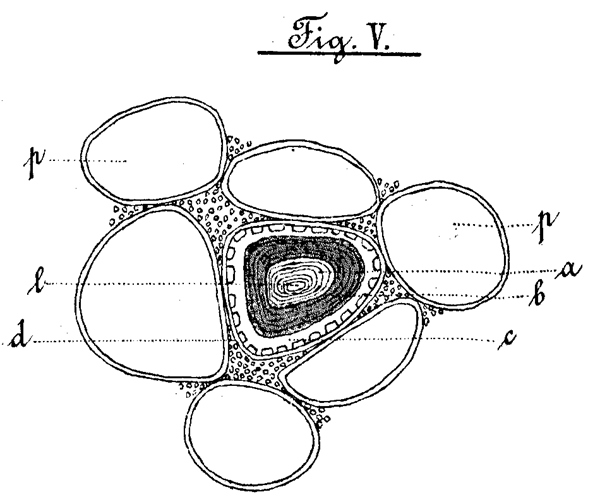
Querschnitt eines Spicularzellenastes des Blattes der Welwitschia mit
den umliegenden Parenchymzellen (vergr. 1:585).
p. Parenchymzellen, l. Lumen.
a. Innenschicht, b. Mittelschicht, c. Krystallschicht, d. Aussenschicht.
Fußnoten
- ↑ Versuch einer allgemeinen physiologischen Chemie pag. 853-855.
- ↑ Grundzüge der wissenschaftlichen Botanik.<
- ↑ Über die physiologische Bedeutung des oxalsauren Kalks.
- ↑ Über Einlagerung von Calciumoxalatkrystallen in die pflanzliche Zellhaut Flor. 1872 Nr. 7, 8, 9.
- ↑ Über Bedeutung der Kalkablagerungen in den Pflanzen, Landwirtschaftl. Jahrbücher 1881.
- ↑ Über Calciumoxalatbildung in den Blättern. Botan. Zeitung 1888.
- ↑ Anatomisch-physiologische Untersuchung der Kalksalze und Kieselsäure in der Pflanze.
- ↑ L'ossalato di calcio nelle foglio Rolletino della società italiana de microscopisti Ann. I. vol. I. Fasc. 1-2 pag. 30-44.
- ↑ Zur Frage der Assimilation der Mineralsalze durch die grüne Pflanze. Flora 1890. Heft 3.
- ↑ On Welwitschia a new genus of Gnotacee by Joseph Dalton Hooker pag 18. 19.
- ↑ On the spiculae contained in the wood of Welwitschia and thee crystals pertaining to them Letter to Dr. Hooker. Journal of the Proc. of the Linnean. Soc. vol. VII.
- ↑ Über einige geformte Vorkommnisse oxalsauren Kalks in lebenden Zellmembranen, Bot. Zeitung 1871, Nr. 33.
- ↑ Die Lehre von der Pflanzenzelle pag. 246.
- ↑ Morphologische Betrachtung der Blätter von Sciadopitys. Bot. Zeit. 1871, pag 8.
- ↑ l. c. p. 8.
- ↑ Anatomie des Gnetacees et des conifères. Annales des sciences naturelles V, Tome XX. pag. 15. 16. 17. Struct. de la feuille, faiseaux médians.
- ↑ l. c. p. 9.
- ↑ Über die Beziehung des anatomischen Baus der Assimilationsorgane zu Klima und Standort mit spec. Berücksichtigung des Spaltöffnungsapparates. Inaug.-Dissert. Freiburg, pag. 11.
- ↑ Vergleichende Anatomie der Vegetationsorgane der Phanerogamen und Farne pag. 313. 338. 348. 388. 425. 434.
- ↑ l. c. p. 9.
- ↑ l. c. p. 6.
- ↑ l. c. p. 9.
- ↑ l. c. p. 6.
- ↑ l. c. p. 8.
- ↑ l. c. p. 8.
- ↑ l. c. p. 6.
- ↑ l. c. p. 8.
- ↑ l. c. p. 9.
- ↑ l. c. p. 13.
- ↑ Ein Beitrag zur Kenntnis der Gefässbündel. Bot. Zeit, 1864, Nr. 22.
- ↑ l. c. p. 9.
- ↑ l. c. p. 13. pag. 347.
- ↑ Über Transfusionsgewebe. Flor. 1880
- ↑ l. c. p. 13. pag. 314.
- ↑ Zur Anatomie von Araucaria brasiliana. Bot. Zeit. 1872 pag. 606. 607.
- ↑ l. c. p. 6. pag. 70.
- ↑ Zur Kenntnis der Einlagerung von Calciumoxalatkrystallen in die Zellmembran. Östr. botan. Zeitschrift 1882, Nr. 12.
- ↑ Untersuchung über Bildung von centrifugalen Wandverdickungen an Pflanzenhaaren und Epidermis. Inaug.-Dissert. Bonn.
- ↑ l. c. p. 3.
- ↑ l. c. p. 6 u. 7.
- ↑ Contribution à l'étude des péricarpes charnus. Du noyau des drupes. Histologie et histogénèse Lyon 1890.
- ↑ Di un processo di tuberifiazione nei tegumenti seminali del genere Tilia L. Atti, della R. Accad. delle scienze. vol. XX, Torino. 1885.
- ↑ Physiologische Chemie der Pflanzen.
- ↑ l. c. p. 6.
- ↑ Fremy, Erdmann, Dragendorff, Schuppe, Koboll, Gruning, Treffner, Koch.
- ↑ Beiträge zur näheren Kenntnis der Holzsubstanz. Sitz-Bericht der Wiener Acad. Bd. 85.
- ↑ Berichte der deutschen chemischen Gesellschaft. Bd. VII. pag. 608. Bd. VIII. pag. 1115. 1123. 1127. 1140. Bd. IX. pag. 52.
- ↑ Histochemische Untersuchungen verholzter Membranen Flor. 1890. Heft I.
- ↑ l. c. p. 41.
- ↑ Die Autooxydation in der lebenden Pflanzenzelle. Bot. Zeit. 1883. Nr. 5 und 6.