Aus dem Max-Planck-Institut für Verhaltensphysiologie in Erling-Andechs. Direktor: Prof. Dr. J. Aschoff
Der Einfluß von Aufwachen und Belichten auf Eosinophilenzahlen, Sauerstoffverbrauch, Pulsfrequenz und Rektaltemperatur des Menschen.
Inaugural-Dissertation zur Erlangung der Doktorwürde
in der gesamten Medizin an der Ludwig-Maximilians-Universität zu München
vorgelegt von Florian Bleisch aus Berlin
Berlin 1978
Mit Genehmigung des Fachbereichs Medizin der Universität München
Berichterstatter: Prof. Dr. rer. nat. R. Wever
Mitberichterstatter: Prof. Dr. med. Thurau
Dekan : Prof. Dr. med. W. Spann
Tag der mündlichen Prüfung: 30.5.1978
Herrn Prof. Dr. R. Wever danke ich für die Überlassung des Themas und für die engagierte Unterstützung bei der Durchführung der Experimente sowie der Auswertung der Ergebnisse.
Mein Dank gilt auch Herrn Prof. Dr. J. Aschoff für die Durchsicht der Arbeit.
Für die immer freundliche Hilfe bei der Lösung mannigfaltiger Probleme in der Durchführung der Arbeit danke ich allen Mitarbeitern des Max-Planck-Institutes für Verhaltensphysiologie in Erling-Andechs.
Inhalt
- Einleitung
- Methodik
- Auswertung
- Ergebnisse
- Diskussion
- Zusammenfassung
- Literaturverzeichnis
- Lebenslauf
Einleitung
Es kann heute als gesichert gelten, daß die bei Tier und Mensch an nahezu allen Organen und in allen Funktionskreisen feststellbaren tagesperiodischen Prozesse ihren Ursprung im Organismus selbst haben, also zur Klasse der endogenen Rhythmen gehören. Die endogenen "circadianen" (Halberg 1960) Rhythmen werden durch periodische Faktoren der Umwelt (Zeitgeber) auf 24 Stunden synchronisiert. Diese Umweltfaktoren beeinflussen häufig, unabhängig von ihrer synchronisierenden Wirkung auf die Rhythmik, einzelne biologische Variable auch unmittelbar in ihrem Verlauf (Maskier-Effekt). Beide Wirkungsweisen können im Tierexperiment für den Faktor "Licht" leicht nachgewiesen werden (Aschoff 1963, 1964). Mit einem Licht-Dunkel-Wechsel lassen sich die circadianen Rhythmen der meisten Tierarten synchronisieren, und zusätzlich wirkt Belichtung auf viele Funktionen im Sinne von Anregung oder auch Hemmung.
Die Frage nach der Wirkung von Licht auf das circadiane System des Menschen kann nicht so eindeutig beantwortet werden. Bei der Synchronisation menschlicher circadianer Rhythmen scheint der Licht-Dunkel-Wechsel nur eine untergeordnete Rolle zu spielen (Wever 1970), und unmittelbare (sofortige) Wirkungen von Belichtung auf einzelne, insbesondere vegetative Funktionen sind nicht häufig belegt. Zu den klassischen Beispielen solcher Wirkung rechnen manche Autoren den morgendlichen Abfall der eosinophilen Leukozyten im Blut. Appel (1939) hat ihn als erster ausdrücklich auf Lichteinfall in die Retina zurückgeführt, und spätere Untersucher sind ihm in dieser Interpretation gefolgt (Appel und Hansen 1952, Levy und Conge 1953, Radnot, Wallner und Honig 1960, Hollwich und Dieckhues 1967).
In jüngerer Zeit haben Hildebrandt und Lowes (1972) aus ihren Messungen der Eosinophilenzahlen, der Herzfrequenz und der Cortisolausscheidung im Harn den Schluß gezogen, daß diese Variablen unmittelbar durch Licht beeinflußt werden, und ferner, daß die Größe dieser Beeinflussung von der Tageszeit abhängt. Die Deutung der Befunde von Hildebrandt und Lowes (1972) ist jedoch ebensowenig zwingend wie die früherer Untersucher, da bei den von ihnen untersuchten Personen der Zeitpunkt des Lichteinfalles mit dem des Aufwachens zusammenfiel. Es läßt sich nicht ausschließen, daß der Wechsel von Schlafen zu Wachen auch ohne Lichteinfall zu Änderungen der Meßgrößen führen kann, wie sie von den Autoren beschrieben worden sind.
Für regelhafte Änderungen vegetativer Funktionen durch den Schlaf sprechen eine Reihe von Befunden. Bei Schlafbeginn vermindert sich die Herzfrequenz (Grützmann 1831, Klewitz 1913, Grollmann 1930, Brooks 1956, Burdick 1970), sinken Sauerstoffverbrauch und Rektaltemperatur (Robin et al. 1968, Th. Schmidt 1972) und steigt die Hauttemperatur (Magnussen 1939, Kleitmann et al. 1948); beim Aufwachen sind die Bewegungen gegenläufig.
Wiederum jedoch ist, wie bei den Untersuchungen über Lichteinflüsse, der Einwand zu machen, daß in den Experimenten zum Nachweis der Schlafwirkung die Retina für die Dauer der Wachzeit belichtet war, daß also Schlaf und Abdunkeln der Retina zusammenfielen. Eindeutige Schlüsse auf die Wirkung von Schlaf oder von Belichtung können aus den Ergebnissen nicht gezogen werden.
Einflüsse der Belichtung lassen sich von solchen des Aufwachens trennen, wenn Versuchspersonen nach dem Wecken für einige Stunden im Dunkeln wach bleiben und erst dann dem Licht ausgesetzt werden. In der hier vorgelegten Arbeit wird über Ergebnisse derartiger Versuche berichtet.
Methodik
Zur Trennung des Einflusses von Aufwachen und Belichten sind Versuche durchgeführt worden, die das abendliche Einschlafen, das morgendliche Aufwachen im Dunkeln, einige Stunden Wachheit im Dunkeln und dann einige Stunden Wachheit im Hellen umfaßten; die Versuche dauerten jeweils mindestens 12 Stunden. Nach sechs vorbereitenden Nächten zur Erprobung der Meßapparaturen sind zwei Serien von Versuchen durchgeführt worden, die sich im Zeitablauf unterscheiden; dabei wurde jede Versuchsperson während zweier Nächte hintereinander untersucht.
Erste Serie: Acht Versuchspersonen wurden untersucht, die zwischen 0.00 und 1.00 Uhr mit dem Schlaf begannen und bis 7.00 Uhr im völligen Dunkel schliefen, dann geweckt wurden, und bis 9.00 Uhr im Dunkeln ruhig wach lagen. Um 9.00 Uhr wurde das Licht angemacht und die Versuchspersonen lagen bis 11.00 Uhr weiter ruhig im Bett, ohne sich in irgendeiner Weise zu betätigen oder zu lesen. Die erste Serie umfaßte also insgesamt 16 Nächte.
Zweite Serie: Vier Versuchspersonen begannen den Versuch ebenso wie die Personen der ersten Serie, lagen aber von 7.00 bis 10.00 Uhr im Dunkeln wach, also drei Stunden lang, um dann weitere zwei Stunden im Hellen wachzuliegen. Die zweite Serie umfaßte also acht Nächte.
Die Versuchspersonen waren zwischen 24 und 30 Jahre alt. Gesunde junge Männer wurden ausgesucht, deren Eosinophilenzahlen gegen Mittag über 50 Zellen/mm3 lagen, da der Zählfehler mit abnehmender Zellzahl größer wird (Lützenkirchen 1951).
Während des Versuches hielten sie sich, bis auf die Unterhose nackt und unzugedeckt, in einer Klimakammer auf, die auf 28,0 + 0,2° C und 50 +/- 5% Luftfeuchtigkeit geregelt
Über eine Gegensprechanlage bestand dauernd die Möglichkeit eines Kontaktes zwischen Versuchsperson und Versuchsleiter, und in der Wachzeit, besonders im Dunkeln, wurden die Probanden laufend ermahnt, wach zu bleiben und die Augen offen zu halten.
Folgende Parameter wurden untersucht :
1. Die Eosinophilen
Die Messung der Eosinophilen war die einzige, die - zur Blutentnahme - die Gegenwart des Versuchsleiters erforderte. Da während des Schlafes der Versuchspersonen kein Blut abgenommen werden konnte, wurden die Eosinophilen um 0.00 Uhr bestimmt und dann ab 7.00 Uhr, d.h. vom Zeitpunkt des Weckens ab, in halbstündigen Intervallen. Um auch im Dunkeln Blut abnehmen zu können, wurde folgende Anordnung getroffen: Während der Blutentnahmen saß der Versuchsleiter in einem innen beleuchteten, lichtdichten Schrank unmittelbar neben dem Versuchsbett, in den die Versuchperson jeweils kurzzeitig ihre Hand durch eine ebenfalls lichtdichte Manschette steckte. Dadurch konnte der Versuchsleiter in üblicher Weise Blut aus der Fingerbeere entnehmen, ohne daß die Versuchsperson - mit Ausnahme ihrer Hand - dem Licht ausgesetzt werden mußte.
Um in die Klimakammer und in den Schrank zu gelangen, verdunkelte der Versuchsleiter den Vorraum der Klimakammer, so daß jede Belichtung des Probanden ausgeschlossen war. Das Blut wurde nach der Dunger'schen Methode (Begemann und Harwerth 1971) verdünnt und gefärbt, drei Minuten auf einem elektrischen Vibrator geschüttelt und sofort ausgezählt.
Die Zellzahl wurde aus dem Zählergebnis von sechs Fuchs-Rosenthal'sehen Zählkammern gemittelt.
2. Die Herzfrequenz
Die Herzfrequenz wurde elektronisch mit einem Herzfrequenzmesser der Firma Siemens aus dem Brustwand-EKG ermittelt und alle zwanzig Sekunden von einem Punktschreiber festgehalten. Leider stand das Meßgerät nur während der ersten Versuchsreihe zur Verfügung.
3. Die Rektaltemperatur
Die Rektaltemperaturwurde mit einem Widerstandsthermometer gemessen und fortlaufend registriert.
4. Der Sauerstoffverbrauch
Der Sauerstoffverbrauch wurde mit dem Metabolie Rate Monitor der Firma Webb Ass., Yellow Springs, USA, ermittelt. Dazu trägt der Proband eine leichte Gesichtsmaske, durch die von einem servogesteuerten Ventilator ein variabler Luftstrom gezogen wird. Eine polarographische Zelle im Ausatmungsluftstrom mißt den Sauerstoffgehalt und regelt den Luftstrom über den Ventilator so, daß die Sauerstoff-Konzentration der abgesaugten Luft konstant bleibt. Die Stärke des Luftstroms und damit - nach Korrektur bezüglich Luftdruck und Temperatur - die Ventilatordrehzahl ist proportional zum Sauerstoffverbrauch.
Der Ventilator war beweglich über dem Bett des Probanden aufgehängt, um so dessen freie Beweglichkeit zu sichern. Vor jedem Versuch wurde die Apparatur mit reinem Stickstoff geeicht, unmittelbar im Anschluß an jeden Versuch wurde die Eichung kontrolliert. Auf Grund zu großer Abweichungen am Morgen mußten drei Versuche von der Auswertung des Sauerstoffverbrauchs ausgeschlossen werden.
Auswertung
Die Zahl der Eosinophilen wurde bei den Kurven einzelner Versuchspersonen in Absolutwerten (Zellen/mm3) angegeben. Zur Mittelung über viele Versuchspersonen wurden nicht di-rekt die Meßwerte benutzt, sondern, zur Ausschaltung der interindividuellen Variabilität, die Abweichung der einzelnen Meßwerte vom Mittelwert über alle Meßwerte des betreffenden Versuchs.
Zur Bestimmung der Herzfrequenz wurden die aufgezeichneten Werte jeweils über 10 Minuten gemittelt. Beim Errechnen der Stundenmittel wurden die 10 Minuten nach den Blutentnahmen ausgelassen, um die frequenzsteigernden Effekte des Schmerzes auszuschließen.
In der gleichen Weise wurde auch der Sauerstoffverbrauch ermittelt.
Die Nachtwerte der Rektaltemperatur bis 6 Uhr früh wurden halbstündlich, die Morgenwerte vom Wecken bis zum Versuchsende alle 10 Minuten aus den Kurven abgelesen.
Ergebnisse
Alle vier gemessenen Parameter unterliegen im Laufe der Versuchszeit systematischen Veränderungen. Im folgenden soll zunächst an Hand von drei individuellen Verläufen gezeigt werden, daß diese Veränderungen bei den verschiedenen Versuchspersonen unterschiedlich sind. Im weiteren sollen dann die über sämtliche Versuchspersonen gemittelten Verläufe betrachtet werden; hierbei zeigen sich - trotz der interindividuellen Unterschiede - Gemeinsamkeiten, die allgemeine Aussagen zulassen.

Abb. 1: Versuchsperson 10: Verläufe von Eosinophilenzahlen , Herzfrequenz, Rektaltemperatur und Sauerstoffverbrauch im ersten (durchgezogene Linien) und zweiten (gestrichelte Linien) Versuch; 0.00 bis 7.00 Uhr Schlaf, um 7.00 Uhr wurde die Versuchsperson im Dunkeln geweckt, um 9.00 Uhr wurde belichtet .
Versuchsperson 10 (Abb. 1) hat in der ersten Nacht durchweg niedrigere Eosinophilenzahlen als in der zweiten Nacht. In beiden Nächten liegen die Aufwachwerte von 7.00 Uhr um 65 Zellen/mm3 höher als die Abendwerte von 0.00 Uhr. Im ersten Versuch schwankt die Eosinophilenzahl nach dem Wecken zunächst unregelmäßig, um dann in einen stetigen Abfall überzugehen; dieser ist zwar nach Einschalten des Lichtes besonders deutlich, setzt aber bereits vorher ein. Die aus den Werten des betreffenden Abschnittes gerechneten Regressionen ergeben für die Dunkelstunden einen Abfall der Eosinophilen um 5,8%/h und für die Lichtstunden einen Abfall um 20,8%/h. Im zweiten Versuch fällt die Eosinophilenzahl nach dem Wecken während der Dunkelzeit stetig ab; nach Einschalten des Lichtes steigt sie zunächst erheblich an, um dann wieder abzusinken, wobei sie aber den Ausgangswert kaum unterschreitet. Die errechneten Regressionen ergeben für die Dunkelstunden einen Abfall von 11,4%/h und für die Lichtstunden von 6,1%/h. Insgesamt zeigen sich in den beiden Versuchen sehr unterschiedliche Bilder. Das im ersten Versuch erhaltene Bild ist vielleicht mit dem des klassischen Eosinophilen-Sturzes vereinbar, das im zweiten Versuch erhaltene jedoch nicht.
Die Pulskurve der ersten Nacht sinkt bis 7.00 Uhr kontinuierlich ab, mit kurzem Zwischenanstieg um 5.40 Uhr, der durch eine Phase von REM - Schlaf oder durch ein Erwachen verursacht sein kann.
Das Minimum der Herzfrequenz liegt bei 43 Schlägen/min um 7.00 Uhr, kurz vor dem Wecken. Durch das Wecken kommt es zu einer erheblichen Pulsbeschleunigung, die aber innerhalb von 20 Minuten wieder zurückgeht. Von nun an ist meist bei den Blutabnahmen eine Beschleunigung festzustellen, die aber innerhalb von 10 Minuten immer wieder zurückgeht. Um 9.00 Uhr, mit dem Lichtanmachen, kommt es dann zu einer weiteren Erhöhung der Frequenz, diesmal aber nicht nur bei den Blutabnahmen, sondern auch dazwischen.
In der zweiten Nacht fällt die Herzfrequenz sehr viel schneller ab als in der ersten. Das Minimum liegt mit 40 Schlägen/ min ganz am Beginn der Nacht, zwischen 1.00 und 1.30 Uhr. Zwischen 2.00 und 4.10 Uhr sowie zwischen 5.20 und 6.40 Uhr ist die Frequenz etwas höher als während der restlichen Zeit der Nacht, was wieder durch Phasen von REM - Schlaf oder durch Erwachen der Versuchsperson bedingt sein kann. Nach dem Wecken zeigt sich ein ähnliches Bild wie in der ersten Nacht mit kurzfristigen Frequenzsteigerungen bei den Blutentnahmen. Im Hellen ist die Frequenz ungefähr so hoch wie in der ersten Nacht.
Insgesamt liegt die Herzfrequenz in der ersten Nacht etwas niedriger als in der zweiten Nacht.
Die Rektaltemperatur ändert sich in der ersten Nacht stark; das Minimum liegt um 4.30 Uhr bei 35,86° C. Abendlicher Abfall wie morgendlicher Anstieg sind gleichmäßig. In der zweiten Nacht ändert sich die Temperatur sehr viel weniger; im ersten Teil dieser Nacht liegt sie unter der Temperatur der ersten Nacht und im zweiten Teil darüber. Nach dem Wecken verläuft die Temperatur in beiden Nächten nahezu gleich.
In Übereinstimmung mit dem Gang der Herzfrequenz erreicht auch die Rektaltemperatur einen kleinen Zwischengipfel um 3.10 Uhr.
Der Sauerstoffverbrauch schwankt stark, so daß eine Beurteilung schwierig ist. In der ersten Nacht ist der Verbrauch zwischen 3.00 und 7.00 Uhr am niedrigsten. Nach dem Wecken kommt es wie bei der Herzfrequenz zu Anstiegen jeweils nach den Blutentnahmen, aber auch zwischen den Entnahmen ist der Verbrauch nicht mehr so niedrig wie im Schlaf. Nach dem Lichtanmachen sind die Anstiege etwas größer als bei Ruhe im Dunkeln.
In der zweiten Nacht ist der Sauerstoffverbrauch - wie die Herzfrequenz - am Anfang der Nacht, zwischen 1.00 und 1.30 Uhr am niedrigsten. Nach dem Wecken verhält sich der Verbrauch wie in der ersten Nacht, nach dem Belichten ist der Anstieg stärker als in der ersten Nacht. Wegen der starken Schwankungen ist der allgemeine Verlauf des Sauerstoffverbrauchs besser in der Darstellung der Stundenmittel (Abb. 2) zu erkennen.

Abb. 2: Versuchsperson 10: Stundenmittel von Pulsfrequenz (oben) und Sauerstoffverbrauch (unten) im ersten (durchgezogene Linien) und zweiten (gestrichelte Linien) Versuch.

Abb. 3: Versuchsperson 10: Herzfrequenz im ersten Versuch zwischen 6.00 Uhr und Versuchsende um 11.00 Uhr, gedruckt mit einer Punktfolgefrequenz von 3 Punkten/min.
Abb. 3 zeigt die Herzfrequenz von Versuchsperson 10 im ersten Versuch zwischen 6.00 und 11.00 Uhr, gedruckt mit einem Punktintervall von 20 Sekunden. Dabei fallen die grossen Unterschiede der Varianz auf, deren Größe nach Burdick et al. (1970) und nach Baust (1971) von der Schlaftiefe und von den Perioden des REM - Schlafes abhängt. Kurz nach 6.00 Uhr wacht die Versuchsperson offenbar einmal auf, schläft dann aber wieder bis zum Wecken um 7.00 Uhr. Dann kommt es zu einem plötzlichen Anstieg von Varianz und Herzfrequenz, doch dieser Anstieg ist nicht von Dauer. Der Proband döst bald wieder, wird dann aber regelmäßig von den Blutentnahmen und den Ermahnungen etwas aktiviert. Die Varianz ist streckenweise recht klein, wird aber in Abständen von 15 - 30 Minuten größer. Aus diesen Beobachtungen läßt sich folgern, daß die Versuchsperson zwischen 7.00 und 9.00 Uhr nicht durchgehend so wach war wie nachher im Hellen.

Abb. 4: Versuchsperson 11: Verläufe von Eosinophilenzahlen Herzfrequenz, Rektaltemperatur und Sauerstoffverbrauch im ersten (durchgezogene Linien) und zweiten (gestrichelte Linien) Versuch; Versuchsablauf wie bei Versuchsperson 10 - Abb. 1
Abb. 4 zeigt als weiteres Beispiel die Verläufe der verschiedenen gemessenen Variablen in den beiden Versuchen eines anderen Probanden. Die Eosinophilenzahlen sind wieder im zweiten Versuch niedriger als im ersten. Im ersten Versuch kommt es nach dem Wecken zu einem Abfall von fast 20% in der ersten Stunde, später wieder zu einem Anstieg; um 9.00 Uhr ist der 7.00 Uhr - Wert wieder erreicht. Nach dem Belichten erfolgt erneut ein kräftiger Abfall von gut 20% in der ersten halben Stunde, wiederum gefolgt von einem Anstieg, der aber in einen Abfall endet.
Nach dem Wecken und nach dem Belichten kommt es hier also zu recht ähnlichen Reaktionen.
Aus den gerechneten Regressionen ergibt sich - im Wachen - für die Dunkelstunden ein Abfall der Eosinophilen um 1,3 und für die Lichtstunden um 10,9%/h. In der zweiten Nacht findet sich nach dem Wecken kaum eine systematische Veränderung; erst nach dem Belichten ist ein deutlicher Abfall festzustellen. Hier ergeben die errechneten Regressionen für die Dunkelstunden einen Abfall der Eosinophilen um 5,5%/h und für die Lichtstunden um 10,8%/h.
Die Herzfrequenz zeigt in beiden Nächten ähnliche Verläufe: Nach dem abendlichen Abfall kommt es zu niedrigsten Werten in der ersten Nacht zwischen 5.20 und 7.00 Uhr, in der zweiten zwischen 5.20 und 5.40 Uhr. Nach dem Wecken geht die Frequenz dann auf ein konstantes Niveau (im zweiten Versuch etwas höher als im ersten), das sich beim Lichtanmachen kaum noch verändert, im zweiten Versuch eher noch etwas abfällt.
Die Rektaltemperatur zeigt in beiden Nächten einen regelmäßigen Abfall und Wiederanstieg, wobei die Werte der zweiten Nacht höher liegen als die der ersten. In der ersten Nacht wurden die niedrigsten Werte um 4.30 Uhr registriert, in der zweiten um 6.20 Uhr. Im Wachen haben, wie bei Versuchsperson 10, beide Kurven annähernd den gleichen Verlauf.

Abb. 5: Versuchsperson 11: Stundenmittel von Pulsfrequenz (oben) und Sauerstoffverbrauch (unten) im ersten (durchgezogene Linien) und zweiten (gestrichelte Linien) Versuch.
Der Sauerstoffverbrauch zeigt in beiden Nächten ungefähr parallele Kurven, doch liegt er in der zweiten Nacht um circa 60 ml/min über dem der ersten. Bis 2.00 Uhr fällt der Sauerstoffverbrauch ab, um 7.00 Uhr mit dem Wecken kommt es in beiden Nächten zu einem starken Anstieg, zu einem weiteren kommt es dann beim Belichten. Klarer zu erkennen sind die Tendenzen auf der Darstellung der Stundenmittel des Sauerstoffverbrauchs (Abb. 5): Die niedrigsten Werte sind in der ersten Nacht zwischen 3.00 und 4.00 Uhr registriert, in der zweiten zwischen 6.00 und 7.00 Uhr. Mit dem Wecken kommt es zu einem kräftigen Anstieg des Verbrauchs um 40 ml/min. In den beiden Dunkelstunden bleibt der Verbrauch konstant, erst mit dem Belichten gibt es wieder einen Anstieg um 40 bzw. 20 ml/min. Aber auch von der ersten zur zweiten hellen Stunde kommt es in beiden Nächten zu einem Anstieg des Verbrauchs um 20 bzw. 50 ml/min.

Abb. 6: Versuchsperson 11: Herzfrequenz im ersten Versuch zwischen 6.00 Uhr und Versuchsende um 11.00 Uhr, gedruckt mit einer Punktfolgefrequenz von 3 Punkten/min.
Die Originalkurve der Herzfrequenz des ersten Versuchs von Versuchsperson 11 (Abb. 6) zeigt, nach einer unruhigen Phase zwischen 6.00 und 6.40 Uhr, beim Wecken um 7.00 Uhr einen Anstieg der Herzfrequenz und ihrer Varianz. Nach anfänglich überschießender Reaktion bleiben dann die Herzfrequenz und Varianz konstant, beim Belichten zeigen sich keine Veränderungen. Dieser Proband ist also nach dem Wecken sofort ganz wach gewesen; das Licht konnte ihn nicht, wie die Versuchsperson 10, noch wacher machen. Nach dem Versuch äußerte er sich auch indigniert zu den häufigen Ermahnungen, wach zu bleiben.
Als letztes individuelles Beispiel werden die Registrierungen der Versuchsperson 14 aus der zweiten Versuchsserie gezeigt (Abb. 7).

Abb. 7: Versuchsperson 14: Verläufe von Eosinophilenzahlen, Rektaltemperatur und Sauerstoffverbrauch im ersten (durchgezogene Linien) und zweiten (gestrichelte Linien) Versuch: 0.00 bis 7.00 Uhr Schlaf, um 7.00 Uhr wurde die Versuchsperson geweckt, um 10.00 Uhr wurde belichtet.
Hier ist besonders der Verlauf der Eosinophilenzahlen von Interesse, da er anders ist als bei den vorher gezeigten Probanden. Diesmal liegen die Werte der zweiten Nacht höher als die der ersten, ansonsten verlaufen beide Kurven weitgehend parallel. Nach dem Wecken fällt die Eosinophilenzahl steil ab, um dann wieder anzusteigen; etwa zwei Stunden nach dem Wecken kommt es erneut zu einem Eosinophilen-Abfall. Auf die Belichtung zeigt sich in keiner der beiden Nächte eine Reaktion. Aus den gerechneten Regressionen ergibt sich für den ersten Versuch ein Abfall der Eosinophilen, der für die drei Dunkelstunden 1,1%/h und für die beiden Lichtstunden 2,3%/h beträgt; für den zweiten Versuch ergibt sich für die Dunkelstunden ein Abfall von 8,8%/h und für die Lichtstunden von 5,8%/h. In allen Fällen schwanken die Verläufe derartig, daß die über mehrere Stunden gemittelten Änderungen wenig besagen. Für die hier anzustellenden Überlegungen scheint es wichtiger zu sein, daß bei dieser Versuchsperson das Aufwachen in beiden Versuchen von einem Eosinophilen-Abfall gefolgt ist, das Einschalten des Lichtes aber eher von einem Eosinophilen-Anstieg.

Abb. 8: Versuchsperson 14: Stundenmittel des Sauerstofferbrauchs im ersten (durchgezogene Linie) und zweiten (gestrichelte Linie) Versuch.
Die Rektaltemperatur ist wieder in der zweiten Nacht höher als in der ersten, das Minimum der ersten Nacht liegt bei 6.20 Uhr, das der zweiten Nacht bei 7.00 Uhr. Der Sauerstoffverbrauch ist in der ersten Nacht deutlich höher als in der zweiten. In beiden Nächten steigt der Verbrauch stärker beim Belichten als beim Wecken. Die Stundenmittel des Sauerstoffverbrauchs (Abb. 8) zeigen eine Erhöhung beim Wecken um 40 bzw. 28 ml/min, beim Belichten um 88 bzw. 84 ml/min.

Abb. 9: Verläufe von Eosinophilenzahlen, Herzfrequenz, Rektaltemperatur und Sauerstoffverbrauch, über alle Versuchspersonen der ersten Serie gemittelt; durchgezogene Linien: jeweils erste Versuche, gestrichelte Linien: jeweils zweite Versuche.
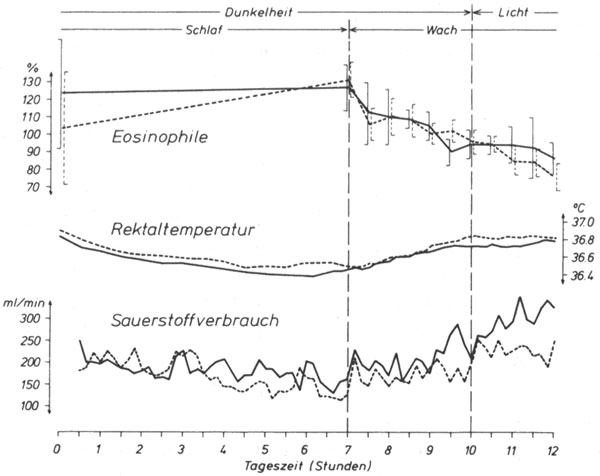
Abb. 10: Verläufe von Eosinophilenzahlen, Rektaltemperatur und Sauerstoffverbrauch, über alle Versuche der zweiten Serie gemittelt; durchgezogene Linien: jeweils erste Versuche, gestrichelte Linien: jeweils zweite Versuche.
Die über alle Versuche je einer Serie gemittelten Kurven sind in den Abbildungen 9 und 10 dargestellt. Im großen und ganzen verlaufen die gemessenen Variablen in den beiden Versuchsreihen ähnlich zueinander. Sauerstoffverbrauch, Rektaltemperatur und Pulszahl (nur Abb. 9) sinken nach dem Einschlafen ab, durchlaufen während der Nacht Minima und steigen nach dem Aufwachen wieder an, ohne daß ein deutlicher Belichtungseffekt zu erkennen ist. Bemerkenswert ist, daß in beiden Versuchsreihen die Rektaltemperatur während der zweiten Versuchsnacht höher war, als in der ersten Nacht. - Besondere Beachtung verdient der Verlauf der Eosinophilenzahlen. Diese stiegen in allen Versuchen während des Schlafes an. Im ersten Versuch der ersten Reihe (Abb. 9) nahmen die Eosinophilen nach dem Wecken im Dunkeln nur wenig ab, im Hellen dann schneller.
Im zweiten Versuch dieser Reihe nahmen sie im Dunkeln zunächst schneller ab, um dann wieder anzusteigen. Nach dem Lichteinschalten (9.00 Uhr) fiel die Eosinophilenzahl erneut ab, um später (10.30 Uhr) ein zweites Maximum zu durchlaufen. Aus diesen Versuchen kann nicht entschieden werden, ob die Lage des ausgeprägten Maximums um 9.00 Uhr durch das Ende der Dunkelstunden gegeben war (möglicherweise haben die Versuchspersonen nach der Erfahrung des ersten Versuchs das Lichteinschalten erwartet): Nach Fortlassen dieses Maximums wäre der Eosinophilen-Abfall während der Lichtstunden nicht stärker als während der Dunkelstunden. Der wellenförmige Verlauf der Eosinophilenzahlen während des zweiten Versuchs war der Anlaß dafür, in einer zweiten Versuchsreihe die Dunkelzeit um eine Stunde zu verlängern In dieser zweiten Versuchsreihe blieben die Versuchspersonen also nach dem Wecken für drei Stunden im Dunkeln. Tatsächlich ist in der zweiten Versuchsreihe (Abb. 10) ein ausgeprägt wellenförmiger Verlauf der Eosinophilenzahlen nicht zu entdecken. Die Eosinophilen fallen in dieser Versuchsreihe nach dem Wecken im Dunkeln erheblich stärker ab als nach dem Lichteinschalten.
In den folgenden Abbildungen sind die erhaltenen Ergebnisse noch einmal für die einzelnen Variablen getrennt dargestellt. Dabei sind für die beiden Versuchsreihen Mittelwerte und Standardabweichungen über jeweils sämtliche Versuchspersonen und die beiden Versuche pro Person aufgetragen.

Abb. 11: Verläufe der Eosinophilenzahlen, gemittelt jeweils über beide Versuche aller Versuchspersonen in der ersten Serie (durchgezogene Linien) und in der zweiten Serie (gestrichelte Linien), mit den zugehörigen Standardabweichungen.
Abb. 11 zeigt den Verlauf der Eosinophilen bei der ersten und zweiten Serie. In beiden Versuchsserien fallen die Eosinophilenzahlen nach dem Aufwachen ab. In der ersten Serie ist der stärkste Abfall in der Zeit unmittelbar nach dem Lichteinschalten. Dieser verstärkte Abfall wird aber wesentlich durch den Wiederanstieg am Ende der Dunkelzeit vorgetäuscht: Läßt man den letzten Dunkelwert außer Betracht, so setzen die Lichtwerte stetig den Verlauf der Dunkelwerte fort. In der zweiten Serie fällt die Eosinophilenzahl am stärksten unmittelbar nach dem Aufwachen. Von diesem Zeitpunkt an bleibt der Abfall regelmäßig, ohne daß nach dem Lichteinschalten eine Änderung des Verlaufs zu erkennen ist. In der ersten Serie fallen die Eosinophilen in den beiden Dunkelstunden um 10,5% bzw. 0,0% ab, in den beiden hellen um 22,5% bzw. 11,5%. Die großen Unterschiede in der stündlichen Abnahme sind bedingt durch den hohen 9-Uhr-Wert, der einen starken Abfall nach dem Belichten vortäuscht. Nach Auslassen dieses Wertes fallen die Eosinophilen ziemlich konstant um ungefähr 10%/h ab. In der zweiten Serie beträgt der Abfall in den Dunkelstunden 11,3%/h, in den Lichtstunden 7%/h. Nach dem Auslassen der ersten zwei Morgenwerte ergibt sich auch hier ein recht konstanter Abfall von 7%/h. Insgesamt fallen also die Eosinophilenzahlen in der ersten Versuchsserie im Hellen schneller ab als im Dunkeln, in der zweiten Versuchsserie dagegen im Dunkeln schneller als im Hellen. Dieser Unterschied zwischen den beiden Serien ist statistisch allerdings nicht zu sichern, da die Reaktionsformen von Versuchsperson zu Versuchsperson stark variieren. Die interindividuellen Unterschiede sind wesentlich größer als die systematischen Unterschiede zwischen den beiden Versuchsreihen.

Abb. 12: Verläufe der Rektaltemperatur, gemittelt jeweils über beide Versuche aller Versuchspersonen in der ersten Serie (durchgezogene Linien) und in der zweiten Serie (gestrichelte Linien), mit den zugehörigen Standardabweichungen.
Bei der Rektaltemperatur (Abb. 12) ist aus den gemittelten Verläufen nicht zu erkennen, daß bei beiden Versuchsreihen die Temperatur während des zweiten Versuches höher liegt als während des ersten Versuches (vgl. Abb. 9 und 10); auf dieses bemerkenswerte Ergebnis wird später noch einmal eingegangen (Abb. 15). Aus Abbildung 12 ist zu erkennen, daß die Rektaltemperatur während der zweiten Versuchsreihe nach dem Einschlafen im Mittel geringfügig niedriger ist als während der ersten Versuchsreihe. Ein Einfluß des Weckens läßt sich weder in den Mittelwertskurven der beiden Versuchsreihen (Abb. 12) noch in den individuellen Verläufen (Abb. 1, 4 und 7) feststellen. Dagegen ist in Abb. 12 deutlich zu erkennen, daß sich der Temperaturverlauf zum Zeitpunkt des Lichteinschaltens abflacht: der morgendliche Anstieg knickt in der ersten Versuchsreihe um 9.00 Uhr und in der zweiten Versuchsreihe um 10.00 Uhr ab.

Abb. 13: Stundenmittel der Herzfrequenz über alle Versuchspersonen der ersten Serie gemittelt, für den ersten Versuch durchgezogene Linien, für den zweiten gestrichelte Linien, mit den dazugehörigen Standardabweichungen.
Die Herzfrequenz konnte nur in der ersten Versuchsserie gemessen werden (Abb. 13). Das Mittel über die acht Versuche liegt wie bei der Rektaltemperatur in der zweiten Nacht höher als in der ersten. Am niedrigsten ist die Herzfrequenz zwischen 5.00 und 6.00 Uhr. Mit dem Wecken kommt es zu einem Anstieg um 1,5 Schläge/min in der ersten Nacht. Beim Belichten sind die Anstiege mit 2,3 bzw. 3,0 Schlägen/min deutlicher. Zwischen den einzelnen dunkeln und den einzelnen hellen Stunden bestehen weit geringere Unterschiede in der Frequenz.

Abb. 14: Stundenmittel des Sauerstoffverbrauchs, gemittelt jeweils über beide Versuche aller Versuchspersonen in der ersten Serie (durchgezogene Linien) und in der zweiten Serie (gestrichelte Linien) mit den zugehörigen Standardabweichungen
Die Stundenmittel des Sauerstoffverbrauchs sind in Abb. 14 gezeigt. Nur in der zweiten Versuchsreihe kommt die nächtliche Abnahme zum Ausdruck; hier ist der Verbrauch zwischen 6.00 und 7.00 Uhr, also unmittelbar vor dem Aufwachen, am niedrigsten. Die morgendlichen Steigerungen bei beiden Versuchsreihen sind eher der Circadianen Periodik als einem Einfluß von Wecken oder Licht zuzuschreiben; die Zunahme beim Wecken oder beim Lichtanmachen unterscheidet sich nicht von der, die zwischen Stunden mit konstanten Bedingungen auftrat, d.h. zwischen zwei hellen oder zwei dunklen Stunden.
Der bemerkenswerte Unterschied der Rektaltemperatur zwischen der ersten und der zweiten Nacht legt Vergleiche auch bei anderen Parametern zwischen den beiden Nächten nahe (Abb. 15).

Abb. 15: Vergleich der jeweils im ersten und zweiten Versuch gemessenen und jeweils über die gesamte Versuchsdauer gemittelten Werte für Herzfrequenz, Rektaltemperatur und Sauerstoffverbrauch bei den einzelnen Versuchspersonen.
Bei der Rektaltemperatur ist der jeweils über die Versuchsdauer gemittelte Wert bei 10 von 12 Versuchspersonen in der zweiten Nacht höher als in der ersten, unmittelbar vorhergehenden. Die statistische Analyse über die aus Abb. 15 zu entnehmenden Differenzen zeigt, daß dieser Unterschied nach dem Zeichentest signifikant ist (p < 0.01). Bei der Herzfrequenz ist das Bild nicht so einheitlich, nur bei 5 von 8 Personen sind die Gesamtmittelwerte der zweiten Nacht höher als die der ersten. Das kann daran liegen, daß in der ersten Nacht einige Personen Schwierigkeiten hatten, einzuschlafen und daher länger eine hohe Herzfrequenz behielten. Bei Versuchsperson 10 ist sowohl Herzfrequenz als auch Rektaltemperatur in der zweiten Nacht niedriger als in der ersten; dieses Verhalten ist dem der meisten Versuchspersonen entgegengesetzt.
Beim Sauerstoffverbrauch ist keine Regelhaftigkeit festzustellen; es gab gleich viel Abfälle wie Anstiege von der ersten zur zweiten Nacht.
Auch bei den Eosinophilen ist das Verhalten unregelmäßig.
7 mal sind die Werte der ersten Nacht höher, 5 mal sind sie niedriger als die der zweiten Nacht. Insgesamt ist daher ein signifikanter Unterschied zwischen den beiden aufeinanderfolgenden Versuchsnächten nur bei der Rektaltemperatur zu beobachten.
Diskussion
1. Eosinophilenzahlen
Den morgendlichen Abfall der Eosinophilen, den Domarus 1931 beschrieb, konnten wir bei allen Versuchspersonen feststellen. Unsere Untersuchungen der Eosinophilen begannen jeweils um 7.00 Uhr früh, da der Schlaf durch Blutentnahmen nicht unterbrochen werden sollte, um so eine sichere Trennung von Schlaf und Wachen zu erreichen. Dadurch läßt sich keine Aussage über das Verhalten der Eosinophilen zwischen 0.00 und 7.00 Uhr machen. Bei der getrennten Untersuchung der Effekte von Wecken und Licht fanden wir, daß sich im Durchschnitt die Steilheiten der Abfälle nicht nennenswert unterscheiden: nach dem Wecken ein Abfall von 11,9%/h, nach dem Belichten 11,8%/h.
Die gefundene Größenordnung des Eosinophilen-Abfalles deckt sich mit der bei anderen Untersuchungen gefundenen. Appel (1939) fand einen morgendlichen Abfall zwischen 7.00 und 10.00 Uhr um 12,4%/h; er machte keine Angaben über die Lichtverhältnisse.
Radnot, Wallner und Honig (1960) fanden bei Licht in dem gleichen Zeitraum einen Abfall um 13,2%/h. Bei einer nicht belichteten Kontrollgruppe sahen sie keinen Abfall, sondern einen Anstieg um ca. 6%/h. Doe (1965) fand, bei dreistündlichen Untersuchungen über 24 Stunden, zwischen 3.00 und 6.00 Uhr 13%/h, zwischen 6.00 und 9.00 Uhr 10%/h Abfall der Eosinophilen. Er machte keine Angaben über die Beschäftigung der Versuchspersonen und das Licht in dieser Zeit; das wahrscheinlichste ist, daß die Versuchspersonen mindestens in der Zeit zwischen 3.00 und 6.00 Uhr schliefen, was bedeuten würde, daß der morgendliche Eosinophilensturz schon sehr früh beginnt und nicht vom Schlaf beeinflußt wird. Die ausführlichen Untersuchungen von Rud (1946) zeigen den morgendlichen Abfall zwischen 6.00 und 10.00 Uhr um 8 bis 12%/h bei nachts arbeitenden Menschen. Beim Schlafenden fand er zwischen 3.30 und 10.00 Uhr einen Abfall um 7%/h. Der Unterschied in den Ergebnissen der vorliegenden Arbeit und denen der oben zitierten Arbeiten von Radnot, Wallner und Honig (1960) und Hildebrandt und Lowes (1972) sowie Fatranska (1968), die alle einen spezifischen Abfall nach dem Belichten angeben, könnte auf einem Unterschied im Wachheitsgrad der Probanden im Dunkeln beruhen. In unseren Versuchen sind die Probanden in der fraglichen Zeit durch ständige Ermahnungen ausdrücklich wach gehalten worden. In den oben zitierten Arbeiten anderer Autoren fehlen Angaben über den Wachheitsgrad der Probanden; Versuchspersonen, die bei völliger Dunkelheit im Bett liegen, haben eine natürliche Tendenz einzuschlafen, wenn sie nicht - wie bei unseren Versuchen - durch ständige Ermahnungen daran gehindert werden. Nach Rud (1946) soll der Schlaf allerdings keinen Einfluß auf den Ablauf der Eosinophilenschwankungen haben.
Bei den einzelnen Individuen haben wir sehr unterschiedliche Reaktionsformen auf das Wecken und das Belichten gefunden. Bei einigen gibt es stärkere Abfälle nach dem Wecken, bei anderen nach dem Belichten; manchmal sind die Abfälle nach dem Wecken und Belichten gleich. Die Eosinophilenzahlen folgen offenbar einem Tagesgang, der durch Aufwachen und Belichten nur geringfügig und interindividuell unterschiedlich modifiziert wird. So hat sicher auch die Auswahl der Versuchspersonen einen großen Einfluß auf die Ergebnisse. Unsere Versuchspersonen wurden zwar ca. alle 10 Minuten angehalten, wachzubleiben und die Augen offen zu halten, doch waren erstens die
Intervalle zwischen den Ermahnungen zu lang, um ein Absinken der Wachheit zwischen den Ermahnungen auszuschliessen, zweitens ist auch bei Personen die nicht schlafen, der Wachheitsgrad unterschiedlich. Möglichenfalls sind interindividuelle Unterschiede in der Wachheit der Probanden während der Dunkelzeit, die trotz der bei allen gleichmäßig erfolgten Ermahnungen noch bestanden haben können, die Ursachen für die beobachteten Unterschiede in den Verläufen der Eosinophilen. Hinweise hierfür bietet der Vergleich der Versuchspersonen 10 und 11 (vgl. Abb. l und 4), bei denen der unterschiedliche Verlauf der Varianzen der Herzfrequenz für unterschiedliche Wachheitsgrade während der Dunkelzeit spricht (vgl. Abb. 3 und 6).
2. Herzfrequenz
Die Abnahme der Herzfrequenz in der Nacht ist seit langem bekannt; Gruetzmann (1831, zit. nach Klewitz 1913) beschrieb sie und gab auch schon ein nächtliches Minimum zwischen 3.00 und 4.00 Uhr an. 1930 hat Grollmann Untersuchungen gemacht, um abzugrenzen, ob die nächtliche Verlangsamung des Herzschlages durch den Schlaf oder durch andere Faktoren bedingt ist. Er zählte den Puls im Schlaf, weckte dann die Versuchsperson, um eine halbe Stunde später noch einmal zu zählen. Der Augenblick des Weckens wurde in verschiedenen Versuchen über die ganze Nacht verschoben, und so kam er zu dem Schluß, daß die Frequenz im Schlaf durchschnittlich um 5 Schläge/min niedriger ist als im Wachen. Außerdem fand er noch einen regelmäßigen Abfall der Frequenz in der Nacht, mit einem Minimum zwischen 2.30 und 6.00 Uhr, und zwar sowohl im Schlaf als auch nach dem Wecken. Wir fanden das Minimum zwischen 5.00 und 6.00 Uhr, später als die oben zitierten Autoren, und auch später als Hildebrandt, der 3.00 Uhr angibt, mit Schwankungen zwischen 1.00 und 6.00 Uhr.
Ober den Lichteinfluß auf die Herzfrequenz sind kaum Untersuchungen bekannt. Hildebrandt und Lowes (1972) fanden ein unterschiedliches Ansteigen der Frequenz nach dem Belichten in Abhängigkeit von der Tageszeit. Allerdings ist dabei unsicher, ob die Probanden im Dunkeln schliefen oder wach waren, und somit, wieweit an dem beobachteten Effekt das Aufwachen beteiligt war. In unseren Untersuchungen ist eine Zunahme der Herzfrequenz sowohl nach dem Wecken als auch nach dem Lichtanmachen festzustellen, wobei die Zunahme nach dem Belichten größer ist als nach dem Wecken. Nur bei zwei Versuchspersonen von acht ergibt sich kei-ne Steigerung nach dem Belichten.
Brooks (1956), Oswald (1962), Snyder (1964) und Burdick (1970) haben Herzfrequenz, Herzfrequenzvarianz und dazu das EEG im Schlaf untersucht und festgestellt, daß sowohl Herzfrequenz wie -varianz von der Schlaftiefe abhängen. Dabei ist die Frequenz weniger verläßlich, da häufig kurzfristige Steigerungen in den folgenden Minuten kompensiert werden, und das Mittel der Frequenz also konstant bleibt.
Die Varianz scheint ein genaueres Bild der Schlaftiefe zu geben, sie nimmt mit der Schlaftiefe ab. Besonders groß ist sie in Perioden des REM - Schlafes. Bei den meisten unserer Versuchspersonen war die Varianz der Herzfrequenz beim Wachen im Dunkeln streckenweise recht klein. Nur bei einigen Versuchen war sie vom Zeitpunkt des Weckens an bis zum Versuchsende immer gleich groß. Daraus ergibt sich, daß bei unseren Probanden "wach" im Dunkeln ein wenig präziser Begriff ist; manche sind hellwach, andere dösen, mit vielen Abstufungen dazwischen. Es ist daher möglich, daß die interindividuell unterschiedlichen Verläufe der Eosinophilenzahlen mit den unterschiedlichen Wachheitsgraden der Versuchspersonen zusammenhängt, daß aber die vorliegenden Befunde nicht die Hypothese stützen, Licht sei der wesentliche Faktor, der diesen Parameter beeinflußt.
3. Rektaltemperatur
Die Rektaltemperatur zeigt den bekannten circadianen Verlauf, die Minima liegen meist kurz vor dem Aufwecken (Aschoff et al. 1974). Die interindividuelle Streuung in der zeitlichen Lage der Minima ist dabei recht groß, sie liegen zwischen 1.30 und 9.20 Uhr, während intraindividuell die Minima nur in einem Fall 3.30 Stunden auseinander lagen; im allgemeinen unterschieden sie sich nur um 1.00 bis 1.30 Stunden.
Die Körpertemperatur liegt bei fast allen Probanden in der zweiten Nacht höher als in der ersten. Diese Tatsache fanden wir nirgends beschrieben; eher wäre in der ersten Nacht, bedingt durch die fremden Versuchsbedingungen, eine Temperatursteigerung im Sinne des "psychogenen Fiebers" (Deutsch, 1926) oder des "Heizaffektes" (Ebbeckr, 1948) zu erwarten gewesen. Die Messung des Sauerstoffverbrauches gibt keinen Anhalt dafür, daß der Stoffwechsel in der ersten Nacht niedriger gewesen wäre als in der zweiten. Andererseits mußten die Versuchspersonen nackt, ohne die gewohnte Bettdecke schlafen, so daß ein verstärkter Wärmeverlust in der ersten Nacht denkbar ist, der schon in der zweiten Nacht durch verstärkte Vasokonstriktion kompensiert wurde.
4. Sauerstoffverbrauch
Unsere Versuche zeigten keine Abhängigkeit des Sauerstoffverbrauchs vom Schlaf oder vom Licht. Das Minimum des Verbrauchs lag kurz vor dem Wecken.
Einige Untersucher fanden im Schlaf einen niedrigeren Sauerstoffverbrauch als im Wachen: Magnussen (1944, zit. nach Kleitmann 1963) um 10%, Robin et al. (1968) um 20%. Grollmann (1930) hat bei seinen Versuchen keinen Unterschied zwischen Schlafen und Wachen gefunden, allerdings stellte er einen nächtlichen Abfall um 15% fest, den mit 12% auch Kreider und Iampietro (1969) beschreiben. Grollmann war der einzige, der den nächtlichen Sauerstoffverbrauch im Schlaf mit dem nächtlichen Verbrauch im Wachen verglichen hat, während Robin normale Tagwerte mit nächtlichen Schlafwerten verglich, die Circadianen Schwankungen des Stoffwechsels also vernachlässigte.
Zusammenfassung
In zwei Serien von Versuchen wurde bei 12 jungen Männern der Verlauf folgender Parameter bei Schlaf im Dunkeln, Wachen im Dunkeln und Wachen im Licht untersucht: Eosinophilenzahlen, Herzfrequenz, Rektaltemperatur und Sauerstoffverbrauch.
Um die Einflüsse getrennt darzustellen wurden in jedem Versuch die drei Zustände nacheinander untersucht:
| Erste Serie | Zweite Serie | |
|---|---|---|
| Schlaf im Dunkeln | 0-7 Uhr | 0-7 Uhr |
| Wachen im Dunkeln | 7-9 Uhr | 7-10 Uhr |
| Wachen im Hellen | 9-11 Uhr | 10-12 Uhr |
Dabei zeigte sich, daß einige Parameter von den verschiedenen Zuständen nicht beeinflußt werden, andere deutliche regelhafte Veränderungen aufweisen und manche in individuell unterschiedlichen Formen reagieren.
In Rektaltemperatur und Sauerstoffverbrauch zeigte sich bei den Probanden keine Reaktion auf die Änderung der Zustände, die tageszeitlichen Schwankungen wurden weder vom Aufwecken noch vom Belichten beeinflußt.
Die Herzfrequenz nahm beim Wecken etwas zu, in stärkerem Maße beim Belichten. Darüber hinaus hat sich gezeigt, daß der Wachheitsgrad, gemessen an der Herzfrequenzvarianz, in individuell unterschiedlichem Maße von der Helligkeit abhängt, das heißt, daß die Versuchspersonen nach dem Wecken im Dunkeln unterschiedlich wach waren.
Die Zahl der Eosinophilen zeigt große interindividuelle Unterschiede in der Reaktion auf Wecken und Belichten: Bei manchen Probanden zeigt sich ein stärkerer Abfall nach dem Wecken und keine Reaktion auf das spätere Belichten, bei anderen zeigt sich ein starker Abfall der Eosinophilenzahl nach dem Belichten. Diese Unterschiede scheinen ebenfalls mit den individuell verschiedenen Wachheitsgraden nach dem Wecken und nach dem Belichten zusammenzuhängen, die sich in der Herzfrequenzvarianz zeigten.
Insgesamt ergibt sich, daß - neben der Tagesperiodik - sowohl das Aufwachen als auch das Belichten unabhängig voneinander morgendliche Änderungen der verschiedenen untersuchten Parameter beeinflussen können.
Literaturverzeichnis
Appel, W.: Über die Tagesschwankungen der Eosinophilen. Zschr. ges. exper. Medizin 104, 15-21, 1939
Aschoff, J.: Comparative Physiology: Diurnal Rhythms. Ann. Rev. Physiol. 25, 521-600 (1963)
Aschoff, J.: Die Tagesperiodik licht- und dunkelaktiver Tiere. Rev. Suisse Zool. 71, 528-558 (1964)
Aschoff, J., Fatranska, M., Gerecke, U., Giedke, H.: Twenty-four-hour rhythms of rektal temperature in humans: Effects of sleep-interruptions and of test-sessions. Pflügers Arch. 346, 215-222 (1974)
Begemann, H., Harwerth, H.-G.: Praktische Hämatologie. Thieme, Stuttgart, 1974 S. 354
Brooks, C. McC. et al. : Sleep and Variations in Certain Functional Activities Accompanying Cyclic Changes in Depth of Sleep. J. appl. Phys. 9, 97-104 (1956)
Burdick, J. A. et al.: Heart-Rate Variability in Sleep and Wakefulness. Cardiology 55, 79-83 (1970)
Deutsch, F.: Das psychogene Fieber. Med. Klinik 32, 1213-1215 (1926)
Doe, R. P. et al.: Relationship of Diurnal Variation in 17-Hydroxycorticosteroid Levels in Blood and Urine to Eosinophils and Elektrolyte Excretion. 3. of Clin. Endocrin. and Metabolism 16, 196-205 (1956)
Domarus, A. v.: Die Bedeutung der Kammerzählung der Eosinophilen für die Klinik. Dtsch. Arch. f. klin. Med. 171, 333- (1931)
Ebbecke, U.: Schüttelfrost in Kälte, Fieber und Affekt Klin. Wochenschr. 26, 609- 613 (1948)
Farner, D. S.: The Testicular Response of White-Crowned Sparrows to Stimulatory Photoperiods in Ahemal Cycles. In: Progress in Photobiology. Herausgeber: Christensen,B.Ch., Buchmann, B.Eisvier, Amsterdam 1961, S. 438 ff.
Fatranska, M.: Einige Bemerkungen zum Einfluß des Lichtes, am Organismus des Menschen. Z. ges. Hyg. 14, 146, 148 (1968)
Grollmann, A.: Physiological Variations During Sleep in the Cardiac Output of Man. Am. 3. of Physiol. 95, 274-284 (1930)
Halberg, F.: Physiologie 24-Hour Periodicity; General and Procedural Considerations with Reference to the Adrenal Cycle. Z. Vitamin-, Hormon- u. Fermentforschung 10, 225 ff (1959)
Hildebrandt, G., Lowes, E. M.: Tagesrhytmische Schwankungen der vegetativen Lichtreaktion beim Menschen. 3. Interdiscipl. Cycle Res. 3, 289-301 (1972)
Hollwich, F., Dieckhues, B. : Augenlicht und Nebennierenrindenfunktion. Deutsche Med. Wochenschr. 92, 2335-2341 (1967)
Kleitmann, N.: Sleep and Wakefulness. Chicago 1963 Kleitmann, N. et al. : Variation in Skin Temperature of the Feet and Hands at the Onset of Sleep. Fed. Proc. 7, 66 (1948)
Klewitz, F.: Der Puls im Schlaf. Dtsch. Arch.f. Klin. Med. 113, 38ff (1913)
Kreider, M. B., Iampietro, P.F. : Oxygen Consumption and Body Temperature during Sleep in Cold Environments. 3. of appl. Physiol. 14, 765-767 (1959)
Lévy, F. M., Conge, G.: Action de la lumiëre sur l'eosinophilie sanguine chez l'homme. Compte rendu Soc. Biol. 147, 586-589 (1953)
Lützenkirchen, E.: Eosinopenische Reaktion und Hypophysen-Nebennierenrinden-System bei Hautkrankheiten. Arch. Dermal. u.Syph. 193, 495-502 (1951)
Magnussen, G.: Vasomotorische Veränderungen in den Extremitäten im Verhältnis zu Schlaf und Schlafbereitschaft. Acta Psychiatr. neurol. 14, 39-54 (1939)
Oswald, I.: Sleeping und Waking. Eisvier, Amsterdam 1962
Radnot, M., Wallner, E., Honig, M.: Die Wirkung des Lichts und des Hydergin auf die Eosinophilen Leukozyten des Blutes. Wiener klin. Wochenschr. 72, 101- 115 (1960)
Robin, E. D. et al.: Alveolar Gas Tension, Pulmonary Ventilation and Blood pH during physiologic Sleep in Normal Subjects. 3. Clin. Invest. 37, 981-989 (1958)
Rud, F.: The Eosinophil Count in Health and Mental Disease. Acta Psychiatr. et Neurol. Suppl. 45 (1946)
Snyder, F. et al.: Changes in Respiration, Heart Rate and Systolic Blood Pressure in Human Sleep. J. Appl. Physiol. 19, 417-422 (1964)
Schmidt, Th.: Thermoregulatorische Größen in Abhängigkeit von Tageszeit und Menstruationszyklus. Med. Diss. München 1972
Wever, R.: Zur Zeitgeber-Stärke eines Licht-Dunkel-Wechsels für die circadiane Periodik des Menschen. Pflügers Arch. 321, 133-142 (1970)
Lebenslauf
Am 12.12.1948 bin ich, Florian Bleisch, in Berlin geboren. Meine Mutter ist die Bühnenbildnerin Hannelore Bleisch, geb. Hoffmann, mein Vater war der Dermatologe Dr. med. Gerhard Bleisch, der 1975 gestorben ist. Meine Geschwister sind: Sebastian, geb. am 25.12.51 und Philine, geb. am 21.3.56.
1974 habe ich die Goldschmiedin Marianne Zschimmer geheiratet, 1975 ist unser Sohn Fabian geboren.
1955 wurde ich in die Grundschule in Berlin-Zehlendorf eingeschult, nach vier Jahren kam ich auf das Französische Gymnasium in Berlin-Reinickendorf, wo ich 1968 das Abitur bestand.
Im gleichen Jahr begann ich das Vorklinische Studium an der Medizinischen Fakultät der Freien Universität Berlin, nach dessen Abschluß ich 1971 die klinische Ausbildung an der Medizinischen Fakultät der Technischen Universität München aufnahm. Am 13.12.1974 bestand ich dort das medizinische Staatsexamen.
Die Medizinalassistentenzeit absolvierte ich 1975/76 in Berlin in folgenden Krankenhäusern: Vier Monate Chirurgie im Behring-Krankenhaus in Berlin-Zehlendorf, vier Monate HNO im Klinikum Steglitz der Freien Universität Berlin, vier Monate innere Medizin im St.-Joseph-Krankenhaus in Berlin-Tempelhof.
Am 30.4.1976 bekam ich die Approbation als Arzt, seit Oktober 1976 arbeite ich auf der Intensivstation der Chirurgischen Abteilung des Auguste-Viktoria-Krankenhauses in Berlin-Schöneberg, (Abt. Priv. Doz. Dr. G. Specht).
Florian Bleisch